研究报告
鲨烯合酶基因的密码子偏性分析 
 作者
作者  通讯作者
通讯作者
计算分子生物学, 2013 年, 第 2 卷, 第 3 篇
收稿日期: 2013年03月21日 接受日期: 2013年03月21日 发表日期: 2013年03月21日
引用格式(中文):
何闪, 张淑杰, 修乐山, 邢朝斌, 2013, 鲨烯合酶基因的密码子偏性分析, 基因组学与应用生物学, 32(2): 232-239
引用格式(英文):
He S., Zhang S.J., Xiu L.S., and Xing Z.B., 2013, Codon Usage Analysis in Squalene Synthase Gene, Jiyinzuxue Yu Yingyong Shengwuxue (Genomics and Applied Biology), 32(2): 232-239
为了分析鲨烯合酶(squalene synthase, SS)基因密码子的使用方式及其影响因素,利用 codon W 和 SPSS 16.0 软件对 47 条来自不同物种的 SS 基因进行多元统计分析、对应性分析。SS 基因密码子 1~3 位碱基 的 GC 含量(GC1, GC2 和 GC3)依次为 51.33%、34.65%和 54.37%,3 个位点的 GC 含量均呈极显著相关关系 (p<0.01),对应性分析的结果表明,第 1 轴显示 30.71%的差异,有效密码子数和 GC3、GC1 和 GC2 的均值与 GC3 之间的相关性均达极显著水平(p<0.01)。筛选出的 26 个最优密码子的第 3 位碱基均为 G 或 C。以 MEGA 5.0 构建的基于 SS 蛋白质序列的进化树比基于 RSCU 的聚类更符合传统的系统发育观点。SS 基因 密码子偏好以 G/C 结尾,使用模式受选择和突变影响,突变对密码子偏好影响较大。
鲨烯合酶(squalene synthase, SS)是生物体内胆固醇合成 、代谢途径中的一种重要的酶(Gu et al., 1998)。在药用植物的次生代谢中,SS 也是三萜皂苷和甾醇生物合成过程中的关键酶,SS 能够催化 2 分子的法尼基焦磷酸(farnesylpyrophosphate, FPP)合成 1 分子的鲨烯(squalene) (Busquets et al., 2008; Kim et al., 2011; Han et al., 2012)。目前已经克隆了包括人(Sum-mersa et al., 1993)、拟南芥(Busquets et al., 2008)、裂殖酵母(Robinson et al., 1993)等在内的众多物种的 SS 基因。通过转基因技术将 SS 转入柴胡(Bupleurum fal- catum)后,能够提高其三萜皂苷和植物甾醇的含量(Kim et al., 2011),通过抑制人参(Panax ginseng) SS 的表达则可降低三萜类的积累量(Han et al., 2012)。
不同物种或是同一物种的不同基因其同义密码子使用是不均衡的,即密码子的使用具有偏性(杨春亮等, 2012)。而密码子偏性的存在,则可对基因的外源表达产生影响,造成外源基因的表达量低下甚至不表达(谢磊等, 2004; 赵武等, 2008),如拟南芥中的 2 个不同构型的 SS 基因在大肠杆菌中进行体外表达,效率较低(Busquets et al., 2008)。根据宿主密码子的偏爱性对外源基因进行了优化,可以在不改变氨基酸序列的基础上提高目的蛋白的表达量,为研究蛋白的纯化和功能做出贡献(Zhou et al., 2012; 唐龙盘等, 2011; 张正平等, 2010)。通过优化第 2、5 和 6 位密码子的 4 个碱基能够明显的提高甘油脱氢酶基因 (glycerol dehydrogenase) 在大肠杆菌中的表达水平(唐龙盘等, 2011)。通过定点突变α-糖基转移酶基因(α-cyclodextrin glycosyltransferase),α- 糖基转移酶产量达到了最高值的 48.9 U/mL (Zhou et al., 2012)。 因此,密码子优化是改善目的基因异源表达水平的一种有效方法。本文通过对 SS 的密码子使用偏性进行分析,构建系统进化树,为下一步开展体外高效表达 SS 蛋白,研究 SS 的生物功能以及调节植物等的次生代谢等提供理论参考。
1结果与分析
1.1各物种SS 基因的 GC 含量和 ENC 值
SS 基因密码子各位置的 GC 含量、编码氨基酸 有效密码子数(effective number of codons, ENC)和密 码子数目的相关关系如表1所示。SS 基因的总体 GC 含量为 46.78%,GC1、GC2 和 GC3 分别为 51.33%、 34.65%和 54.37%。整个编码区的 GC 含量与 GC1、 GC2、GC3 之间的相关性均达到了极显著水平(p<0.01)。GC1、GC2 与 GC3 之间相关性达到极显著水 平(p<0.01),且表现为 GC3>GC1>GC2,说明 SS 基因密码子的第 1、2、3 位碱基的组成较相似(r 分别为 0.626, 0.853 和 0.555),不存在显著差异(表2)。由表2中 GC 与 GC1、GC2 和 GC3 之间的相关系数(r 分别为 0.912, 0.656 和 0.987)可知 GC 与 GC1、GC2 之间的相关性更强。ENC 的最小值为 32.30,最大值为 61,均值为 51.03,ENC 值均比较大,密码子存在偏性但不明显,且 ENC 值和密码子第 1、2 和 3 位的碱基的相关性未达到显著水平(表2)。
|
表1各物种 SS 基因编码区各部位的 GC 含量及 ENC 值 |
.gif)
|
表2 SS 基因各相关参数的相关性分析 |

1.2各物种 SS 基因密码子的偏性
1.2.1 ENC 和 GC3 的相关性分析
以 47 个物种 SS 的GC3 值和 ENC 值分别为横、纵坐标绘图(图1)。由图可知,SS 沿期望值曲线分布或落在期望值曲线的下方,但大多数距离期望值曲线较近,只有少数远离期望值曲线。大多数的 SS 基因偏性较弱,少数基因偏性较强。说明 SS 的密码子偏性形成受突变和选择的共同作用,其中碱基突变的影响更强烈。
|
图1 ENC 和 GC3 的相关性分析 |
.gif)
1.2.2各物种 GC12 和 GC3 的关系
各物种 SS 基因的中性绘图分析结果如图2所 示,多数 SS 基因主要分布于对角线下方。GC3 的分布范围在 0.131 4~0.817 4 之间,GC12 的分布范围在0.358 4~0.506 5 之间,GC12 分布较为紧凑。GC3 分布范围较大,主要是由于白色念珠菌(Candida albi- cans)的 GC3 值(GC3 为 0.131 4, GC12 为 0.366 4)偏小引起的。GC12 与 GC3 的相关性双尾检验达到极显著水平(p <0.01),回归曲线斜率为 0.149 7,说 明 GC12 与 GC3 的相关性很强,密码子 3 个位置的碱基组成无差异,密码子的偏好受突变的影响,选择在密码子偏性形成过程中作用较弱,突变在此过程中的作用较强。
|
图2中性绘图分析 |
.gif)
1.3对应性分析
以同义密码子相对使用频率( relative synony- mous codon usage, RSCU)为基础,进行对应性分析,第 1 轴显示了 30.71%的差异,第 2、3、4 轴分别显示 了 13.52%、10.80%和 9.40%的差异。前 4 轴差异合计 64.43%,以第 1 轴为主,其余各轴差异不明显。第 1 轴与 GC3 双尾检验达到极显著水平(p<0.01),呈极显著正相关关系(r=0.989)。第1轴与密码子适应指数(codon adaptation index, CAI) 和密码子偏好指数(codon bias index, CBI)的相关性达到双尾侧检验极显著水平(p<0.01, r 分别为 0.758 和 0.766)。说明GC3 对 SS 基因密码子的偏性有较大的影响,同时物种差别等其它因素对 SS 基因密码子的偏性也有一定影响,其使用模式形成过程较为复杂。
根据各物种的进化水平,将 SS 基因分为:微生物类的 SS 基因(编号 1~20, 共 20 条)、动物类的 SS基因(编号 21~26, 共 6 条)和植物类的 SS 基因(编号 27~47, 共 21 条)。以第 1 轴为横坐标,第 2 轴为纵坐标将各物种 SS 基因分布于平面中。除了动物中的 Naegleria gruberi 的 SS 基因,微生物中的白色念珠菌和酿酒酵母(Saccharomyces cerevisiae) 的 SS 基因相对分散度较大外,其它的动物、植物和微生物的 SS 基因则集中分布(图3)。这说明动物、植物、微生物类 的各物种 SS 基因的碱基构成更为相似,SS 基因各自分布更为集中。
|
图3基于 RSCU 的对应性分析 |
.gif)
1.4最优密码子分析
通过比较 47 条不同物种的 SS 基因,根据其 ENC 值的大小,筛选高、低表达基因,高表达基因的 ENC 均值为 49.59,低表达基因的 ENC 均值为 46.82,分别计算高表达和低表达基因中各密码子的 RSCU 值, 确定 了 SS 基因中存在除了 AUG、 GUG、UAG、UGG、AGG、GGG 以外的 26 个以 G/C 为第 3 位的密码子为最优密码子。SS 基因最优密码子使用频率(frequency of optimal codons index, FOP) 的范围为 0.31~0.54 之间 ,均值为 0.41,最优密码子的使用频率较高。SS 基因偏好使用 G/C 结尾的密码子。
1.5基于 SS 基因密码子偏性的聚类分析
在对基因密码子使用频率进行聚类分析的过程中,将每一条基因作为一个对象,将密码子的 RSCU 值作为变量,利用 SPSS 16.0 进行聚类分析,聚类结果见图4A。由 MEGA 5.0 的 neighbor-joining 连接法构建的 47 条 SS 蛋白的系统发育树如图4B 所示,20 种微生物首先聚为一支,动植物和原虫的亲缘关系较近聚为一支,随后和微生物聚为一个大的分支。通过比较系统发育树的结果可知,在 2 种不同方法构建的系统发育树中,动物物种的 5 条 SS 基因都聚为 一支,且聚类结果相同;植物类和微生物类物种的 SS 基因大体上聚类结果一致。但是基于 RSCU 的聚类结果显示,微生物类物种中的 Ajellomyces capsulatus 和 A. dermatitidis 的 SS 基因和植物类玉米聚为一 类,而基于氨基酸序列的聚类结果则显示,上述 2 个物种与其它微生物类物种聚为一个大类,随后和动植物聚为一类。另外,在依据 RSCU 进行的聚类分析中,N. gruberi 和白色念珠菌聚为一类,和其它的物种距离均较远,而在基于氨基酸序列的进化树中则和植物聚为一类。
|
图4 SS 基因构建的系统发育树 |
.gif)
2 讨论
大肠杆菌中的稀有密码子包括 AGA、AGC、 AUA、CCG、CCU、CUC、CGA 和 GUC,共 8 个(Gros- jean and Fiers, 1982),若外源基因中包含这些稀有密码子时,会显著降低其表达效率(谢磊等, 2004)。SS 基因的所有密码子中 ,RSCU >1.0 的 CUC、GUC、 CCU、CGA、AGC、AGA (RSCU 分 别 为 1.35, 1.17, 1.08, 1.01, 1.04 和 1.29),这些密码子属于大肠杆菌表达系统的稀有密码子,因此利用大肠杆菌作为受体进行原核外源表达 SS 基因时,应将其优化为大肠杆菌的常用密码子,以提高外源基因的表达量。在以酵母为受体菌,进行真核外源表达 SS 基因时,同样应该根据酵母的密码子使用频率分布信息(http: //www.kazusa.or.jp/codon),对外源基因的对应密码子进行优化才能获得良好的体外表达。这种改造外源基因的密码子以消除其存在的宿主中的稀有密码子,而提高表达效率的研究,已在其它基因中被证实。如通过定点突变扩展青霉(Penicillum expansum) 脂肪酶基因中毕赤酵母的稀有密码子后,目的蛋白的表达量比野生基因重组子提高了2.3~2.5 倍(张正平等, 2010)。因此,根据对 SS 基因密码子偏性分析的结果,结合宿主密码子偏性的特点进行优化后,对提高 SS 基因的外源表达效率也同样适用。
生物体内密码子偏性的存在是选择、突变和遗传漂变相互平衡的结果(Bulmer, 1987; 晁岳恩等, 2011)。密码子的使用并不是随机的,不同基因的密码子使用偏性存在不同的特点(Guo et al., 2012)。偏爱使用以 A 或 T 碱基结尾的密码子的基因有双子叶植物 waxy 基因(刘汉梅等, 2010),重要的抗寒调控转录因子 ICE1 (时慧等, 2012)等。偏爱使用以 G 或 C 碱基结尾的密码子的有单子叶植物的 waxy 基因(刘汉梅等, 2010)和大豆光周期抑制开花的重要抑制因子 RAV (related to ABI3/VP1) (杨春亮等, 2012)。SS 基因密码子分析结果显示了 SS 基因偏好于使用以 G 或 C 碱基结尾的密码子,与上述的 waxy 和 RAV 基因的密码子偏性较为相似。SS 基因在突变、选择等各种因素的作用下,产生了密码子偏性,其中突变的作用比选择的作用更强,这符合单基因密码子偏性形成的原因,多数是突变作用大于选择作用的特点(刘汉梅等, 2010)。
以不同物种 SS 基因的 RSCU 为基础构建的系统进化树和基于 SS 蛋白差异构建的系统发育树分 析结果的总体趋势一致,但不完全相同。这种基于 RSCU 与蛋白序列不完全相同的现象在其它研究中也出现过(时慧等, 2012; 晁岳恩等, 2011)。造成聚类 分析结果有偏差的原因可能是由于单个基因在物种中的突变,使密码子偏性出现大的不同,通过增加分析的基因类型和聚类时使用更多的指标,来弥补分析单一因素聚类的不足。基于 RSCU 的聚类分析可以作为传统的进化树建树的参考。
鲨烯合酶在初级代谢和次级代谢中均扮演着重要的角色。其催化合成的三萜皂苷类往往是有生理、药理活性的物质。本研究通过对 SS 基因密码子偏性进行分析,明确了不同物种中 SS 密码子使用的共同特点,为 SS 基因进行密码子优化,规避使用宿主稀有密码子和提高体外表达的效率提供借鉴。
3 材料和方法
3.1目的基因的获取和软件
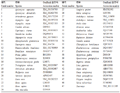
本研究所采用的各物种 SS 基因的全长编码区序列(coding DNA sequence, CDS)信息均来自 GenBank。参照文献(时慧等, 2012)进行密码子分析时采用的 CDS 筛选原则,筛选符合条件的 SS 基因。根据该标准,共获得了 47 条来自不同物种的 SS 基因作为本研究的分析样本(表3)。
|
表3 SS 基因完整编码区核苷酸序列的来源 |
使用在线分析软件 codon W(http://mobyle.pas teur.fr/cgi-bin/portal.py?#forms::codonw)和 CUSP(http:// mobyle.pasteur.fr/cgi-bin/portal.py?#forms::cusp),统 计 GC1、GC2、GC3 和 GC 值等指标,同时统计 CAI、 RSCU、ENC、CBI 和 FOP 等值。
3.2同义密码子偏性分析
通过 codon W 1.4.4 在线软件计算样本的总体 GC 含量,记为 GC。同时利用该软件计算 RSCU、ENC、CAI、CBI 和 FOP,并进行对应性分析。利用 CUSP 软件分别计算密码子第 1、2 和 3 位碱基的 GC 含量,分别记作 GC1、GC2、GC3。使用 SPSS 16.0 软件分析各变量之间的相关性。
ENC 值是目前公认在评价基因整体密码子偏性 用法中最具有参考价值的参数,该值用于衡量某条 基因或某个物种的密码子偏好程度,其取值为 20~61,密码子偏性越大,该值越小,反之亦然(时慧等, 2012)。GC 测量的是基因中 G 和 C 的含量。GC3 则计算密码子第三个碱基中出现 G 或 C 的频率。
3.3 ENC 与 GC3 的相关性分析
利用 ENC 和 GC3 值构建散点图。同时构建标准曲线,即密码子偏性仅由碱基组成决定时的基因 预 期 值 ,计算公式为:ENC = 2 + GC3 + 29/ [ GC32 + (1-GC3)2]。ENC 和 GC3 的相关关系可以用来分析密码子的使用受到突变和选择影响的幅度。当某基因的密码子偏性仅受突变影响时则分布于曲线附近,若其受选择影响较大则分布于曲线下方较远的位置(尚明照等, 2011)。
3.4中性绘图分析
将各物种 SS 基因的 GC12 (GC1 和 GC2 的平均值)作为纵坐标,GC3 作为横坐标进行绘图,分析 SS 基因密码子的 3 个位点碱基组成之间的相关性, 确定影响密码子偏性的因素。如果 GC3 与 GC12 相关性显著,说明 3 个位点的碱基组成相同,碱基突变在密码子偏性的形成中影响较大。否则,基因组 GC 含量高度保守,密码子不同位点的碱基组成不同,密码子的偏好特点受选择的影响较大。
3.5最优密码子分析
将 SS 基因中具最高和最低 ENC 值的各 10% 样本分别作为高、低表达组。计算高、低表达组的RSCU 差值,两者间的 RSCU 差值>0.08 的密码子确定为高 表达最优密码子(尚明照等, 2011)。
3.6各物种 SS 基因的聚类分析
利用 SPSS 16.0 软件分析各条 SS 基因的 RSCU 值,进行基于密码子使用偏性的聚类分析。以密码子 为分组,采用 59 个密码子(去除编码蛋氨酸的密码子 AUG 和编码色氨酸的密码子 UGG 以及 3 个终止密 码子 UAA, UAG, UGA)的 RSCU 值。将各条 SS 基因 的 RSCU 值构建数据库,对密码子使用概率分析时, 将每一条基因作为一个对象,RSCU 作为变量,利用 各密码子 RSCU 值的欧式距离平方测定组间距离, 把最接近的 2 类合并为一个新类,最后分为 n-1 类, 输出树状结构图。
使用 MEGA 5.0 软件,采用 neighbor-joining 连 接法,构建 47 条 SS 基因编码蛋白质的氨基酸序列 的系统进化树。
作者贡献
本文在邢朝斌副教授的指导下,由何闪完成研究的设计、实施、数据整理以及论文撰写,修乐山进行数据汇总和整理,经由张淑杰教授共同修改后完成。
致谢
本研究由国家自然科学基金(30701086)、河北省自然科学基金(C2009001252)和河北省自然科学基金-石药集团医药联合研究基金(H2012401006)共同资助。在实验过程中得到了龙月红老师和吴鹏老师的大力帮助和支持,也得到了河北联合大学生命科学学院周秘、柴丽花、朱金丽、李宝财等同学的有益建议和帮助,在此,一并表示衷心的感谢。
Bulmer M., 1987, Coevolution of codon usage and transfer RNA abundance, Nature, 325(6106): 728-730
Busquets A., Keim V., Closa M., Arco A.D., Boronat A., Arro M., and Ferrer A., 2008, Arabidopsis thaliana contains a single gene encoding squalene synthase, Plant Mol. Biol., 67(1-2): 25-36
Chao Y.E., Chang Y., Wang M.F., He S.L., Zhao X.L., and Lei Z. S., 2012, Codon usage bias and cluster analysis on chloroplas-tic genes from seven crop species, Huabei Nongxuebao (Acta Agriculture Boreali-Sinica), 27(4): 60-64 (晁岳恩, 常阳, 王 美芳, 何盛莲, 赵献林, 雷振生, 2012, 7 种作物叶绿体基因的 密码子偏好性及聚类分析, 华北农学报, 27(4): 60 -64)
Chao Y.E., Wu Z.Q., Yang H.M., He N., and Yang P., 2011, Cluster analysis and codon usage bias studies on psbA genes from 11 plant species, Henong Xuebao (Journal of Nuclear Agricultural Sciences), 25(5): 927-932 (晁岳恩, 吴政卿, 杨 会民, 何宁, 杨攀, 2011, 11 种植物 psbA 基因的密码子偏 好性及聚类分析, 核农学报, 25(5): 927-932)
Grosjean H., and Fiers W., 1982, Preferential codon usage in prokaryotic genes: The optimal codon-anticodon interaction energy and the selective codon usage in efficiently expressed genes, Gene, 18(3): 199-209
Gu P., Ishii Y., Spencer T.A., and Shechter I., 1998, Func- tion-structure studies and identification of three enzyme do- mains involved in the catalytic activity in rat hepatic squa- lene synthase, The Journal of Biological Chemistry, 273 (20): 12515-12525
Guo F.B., Ye Y.N., Zhao H.L., Lin D., and Wei W., 2012, Uni- versal pattern and diverse strengths of successive synony- mous codon bias in three domains of life, particularly among prokaryotic genomes, DNA Research, 19(6): 477-485
Han J.Y., In J.G., Kwon Y.S., and Choi Y.E., 2012, Regulation of ginsenoside and phytosterol biosynthesis by RNA interfer- ences of squalene epoxidase gene in Panax ginseng, Phyto- chemistry, 71(1): 36-46
Kim Y.S., Cho J.H., Park S., Cho J.H., Han J.Y., Back K., and Choi Y.E., 2011, Gene regulation patterns in triterpene biosynthetic pathway driven by overexpression of squalene synthase and methyl jasmonate elicitation in Bupleurum fal- catum, Planta, 233(2): 343-355
Liu H.M., Zhao Y., Gu Y., Zhang H.Y., and Huang Y.B., 2010, Characterization of codon usage of waxy genes in several plants, Henong Xuebao ( Journal of Nuclear Agricultural Sciences), 24(3): 476-481 (刘汉梅, 赵耀, 顾勇, 张怀渝, 黄 玉碧, 2010, 几种植物 waxy 基因的密码子用法特性分析, 核农学报, 24(3): 476-481)
Robinson G.W., Tsay Y.H., Kienzle B.K., Smith-Monroy C.A., and Bishop R.W., 1993, Conservation between human and fungal squalene synthetases: Similarities in structure, func- tion, and regulation, Mol. Cell Biol., 13(5): 2706-2717
Shang M.Z., Liu F., Hua J.P., and Wang K.B., 2011, Analysis on codon usage of chloroplast genome of Gossypium hirsutum, Zhongguo Nongye Kexue (Scientia Agricultura Sinica), 44 (2): 245-253 (尚明照, 刘方, 华金平, 王坤波, 2011, 陆地棉 叶绿体基因组密码子使用偏性的分析, 中国农业科学, 44 (2): 245-253)
Shi H., Wang Y., Yang L.C., and Ding Z.T., 2012, Analysis of codon bias of the cold regulated transcription factor ICE1 in tea plant, Yuanyi Xuebao (Acta Horticulturae Sinica), 39(7): 1341-1352 (时慧, 王玉, 杨路成, 丁兆堂, 2012, 茶树抗寒 调控转录因子 ICE1 密码子偏性分析, 园艺学报, 39(7): 1341-1352)
Summersa C., Karstb F., and Charles A.D., 1993, Cloning, ex- pression and characterisation of the cDNA encoding human hepatic squalene synthase, and its relationship to phytoene synthase, Gene, 136(1-2): 185-192
Tang L.P., Yu J.C., Dai D.F., and Fang B.S., 2011, Expression of glycerol dehydrogenase gene in Escherichia coli by codonoptimization, Weishengwu Xuebao (Acta Microbiologica Sinica) 51(4): 504-509 (唐龙盘, 余劲聪, 戴丹凤, 方柏山, 2011, 甘油脱氢酶基因在大肠杆菌中的密码子优化表达, 微生物学报, 51(4): 504-509)
Xie L., Sun J.B., Zhang S.Q., and Huang J.S., 2004, Research progress in the E. coli expression system, Huanan Redai Nongye Daxue Xuebao (Journal of South China University of Tropical Agriculture), 10(2): 16-20 (谢磊, 孙建波, 张世 清, 黄俊生, 2004, 大肠杆菌表达系统及其研究进展, 华南 热带农业大学学报, 10(2): 16-20)
Yang C.L., Wang L., Wu B., Zhao L., Zhang X.L., and Zhong S. Q., 2012, Codon bias analysis of soybean GmRAV gene, Dongbei Nongye Daxue Xuebao (Journal of Northeast Agri- cultural University), 43(7): 34-41 (杨春亮, 王良, 武斌, 赵琳, 张晓丽, 钟淑琦, 2012, 大豆 GmRAV 基因的密码子偏好性分析, 东北农业大学学报, 43(7): 34-41)
Zhang Z.P., Yang J.K., Xu L., Liu Y., and Yan Y.J., 2010, Cloning, codon optimization and expression of mature lipase gene Penicillum expansum, Weishengwu Xuebao (Acta Mi- crobiologica Sinica), 50(2): 228-235 (张正平, 杨江科, 徐 莉, 刘云, 闫云君, 2010, 扩展青霉脂肪酶基因克隆、密码子优化及表达, 微生物学报, 50(2): 228-235)
Zhao W., Li B., Liang J.X., Liang B.Z., Yao R.Y., Huang A.G., He Y., and Jiang Y.W., 2008, Codon preference analysis of principal non-structural and structural proteins of porcine parvovirus, Guangxi Nongye Shengwu Kexue (Journal of Guangxi Agriculture and Biology Science), 27(4): 299-303, 314 (赵武, 李斌, 梁家幸, 梁保忠, 姚瑞英, 黄安国, 何颖, 蒋玉雯, 2008, 猪细小病毒主要非结构蛋白与结构蛋白的密码子偏爱性分析, 广西农业生物科学, 27(4): 299-303, 314)
Zhou J., Liu H., Du G., Li J., and Chen J., 2012, Production of al- pha-cyclodextrin glycosyltransferase in Bacillus megaterium MS941 by systematic codon usage optimization, J. Agric. Food Chem., 41(60): 10285-10292