研究报告/Research Report
苦荞转录因子基因FtbHLH3的克隆及其非生物胁迫下的表达分析 
2西昌学院, 西昌, 615000
 作者
作者  通讯作者
通讯作者
计算分子生物学, 2016 年, 第 5 卷, 第 2 篇
收稿日期: 2016年03月10日 接受日期: 2016年03月10日 发表日期: 2016年03月10日
引用格式(中文):
姚攀锋, 赵学荣, 李茂菲, 王安虎, 吴琦, 2016, 苦荞转录因子基因FtbHLH3的克隆及其非生物胁迫下的表达分析, 基因组学与应用生物学, 35(2): 429-435 (doi: 10.13417/j.gab.035.000429)
引用格式(英文):
Yao P.F., Zhao X.R., Li M.F., Wang A.H., and Wu Q., 2016, Cloning and Expression Analysis of Transcription Factor Gene FtbHLH3 from Fagopyrum tataricum under Abiotic Stress, Jiyinzuxue Yu Yingyong Shengwuxue (Genomics and Applied Biology), 35(2): 429-435 (doi: 10.13417/j.gab.035.000429)
根据苦荞花期转录组数据,采用RT-PCR技术和PCR技术克隆得到一条新的bHLH类蛋白基因FtbHLH3 (登录号: KU296217),并分析了逆境胁迫下该基因在芽期苦荞胚轴和子叶中的表达量。结果显示,FtbHLH3基因cDNA全长1 273 bp,包含一个957 bp的开放阅读框,编码蛋白含318个氨基酸。蛋白结构域分析表明,FtbHLH3编码蛋白N-端和C-端分别含有一个bHLH家族典型的结构域。系统进化树分析表明,FtbHLH3与其它植物参与抗逆的bHLH蛋白聚为一簇。荧光定量PCR分析表明,UV-B处理苦荞后,胚轴中FtbHLH3相对表达量在6 h内缓慢增加至1.08,12 h后显著上升至48 h的2.96;而子叶中FtbHLH3相对表达量在3 h后就显著上升,至24 h达到最大值6.64后趋于稳定。4℃冷处理苦荞后,胚轴和子叶中FtbHLH3相对表达量均随时间持续上升,并在48 h后达到最大值,分别为3.22和10.27。可见,本研究克隆的FtbHLH3基因可能参与了苦荞对UV-B和寒冷等非生物胁迫的应答反应。
苦荞(Fagopyrum tatarium)也称鞑靼荞麦,属蓼科植物,原产于我国及印度等地。由于抗逆性强,其主要分布在四川、云南、贵州和西藏等西部省区的一些干旱高寒山区、少数民族地区和边远山区。随着对苦荞营养价值和保健功能的深入研究,对苦荞的利用和开发也越来越受到重视(Kim et al., 2001)。具有均衡营养结构和药食两用的苦荞逐渐作为一种健康食品被加工为各种面食、糕点和调味品等,受到消费者的广泛青睐(Bonafaccia, 2003)。
非生物胁迫是当前植物生长以及粮食产出的主要制约因素之一(Zhou et al., 2015)。干旱、UV-B、极端温度是导致农作物品质和产量下降的主要非生物因素。植物在遭受非生物胁迫后通过体内生理生化和分子机制调节,从而快速修复逆境胁迫所造成的损伤,以此适应不断变化的环境(Sha et al., 2015)。在众多调控机制中,转录因子所介导的环境适应性变化至关重要,如MYB、HLH和WRKY等转录因子都广泛参与调控与干旱、UV-B、极端温度、激素和病原反应等相关基因的表达(Li et al., 2015)。
bHLH转录因子在多种植物中广泛存在,是构成真核生物蛋白的一个大家族,因含有bHLH结构域而得名。研究证明该家族具有特有的碱性区域/螺旋-环-螺旋基序(bHLH,basic helix-loop-helix),其中碱性区域能识别靶基因启动子区域中顺式作用元件E-box (5'-CANNTG-3')核心序列,与DNA结合相关;而HLH结构域是同其他相关蛋白形成杂合二聚体所需的(周华等, 2015)。
近年来,已经从拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)等多种植物中分离、克隆出bHLH基因(Ding et al., 2009; Salvatierra et al., 2013),并对其调控功能进行了研究,发现bHLH在植物生长发育和形态建成、胁迫响应以及植物次生代谢中发挥着重要作用(Ding et al., 2009; Matus et al., 2010)。目前,有关苦荞转录因子的研究主要集中在参与类黄酮代谢(Bai et al., 2014)和非生物胁迫应答(Zhou et al., 2011; 黄云吉等, 2015)的MYB转录因子家族,进一步挖掘参与苦荞抗逆的bHLH等其他转录因子可加深对苦荞抗逆机制的理解。本实验根据获得的苦荞花期转录组数据,采用RT-PCR技术克隆苦荞抗逆相关的FtbHLH3基因,探究FtbHLH3基因的表达对UV-B和4℃冷胁迫处理的应答关系,为其进一步的功能鉴定奠定基础。
1结果与分析
1.1苦荞FtbHLH3基因cDNA序列的克隆
以苦荞cDNA为模版,采用特异引物FtbHLH3f和FtbHLH3r,PCR扩增得到一条大小约为1 kb的特异条带(图1)。测序结果表明该片段大小为951 bp,编码316个氨基酸残基的蛋白FtbHLH3 (图2)。经过NCBI中的Pro-BLAST比对显示:该蛋白与其它植物bHLH的同源性在46%~68%之间,表明克隆得到的序列属于类bHLH基因家族。将该基因命名为FtbHLH3,其在GenBank中的注册号为:KU296217。
|
图 1 苦荞FtbHLH3 基因的扩增
Figure 1 Amplification of FtbHLH3 gene from F. tataricum
|
.png)
|
图 2 FtbHLH3 核苷酸序列及其推导的氨基酸序列
Figure 2 Nucleotide and deduced amino acid sequence of FtbHLH3
|
.png)
1.2编码蛋白FtbHLH3的理化性质预测
用ExPASy-ProtParam 对编码蛋白FtbHLH3的理化性质分析表明,其编码蛋白分子式为C1588H2517 N425O503S16,相对分子质量为36.12 kD,理论pI值为4.89其中有37个氨基酸残基(Arg+Lys)带正电荷,49个氨基酸残基(Asp+Glu)带负电荷;不稳定系数为78.77,是不稳定性蛋白(<40为稳定性蛋白),平均亲水性数为-0.488。Singal P的信号肽预测显示,该蛋白序列无信号肽位点,属于非分泌性蛋白质。Tmpred的跨膜结构域预测分析表明,该蛋白为跨膜蛋白。Target P软件预测得知FtbHLH3的mTP值为0.050,SP值为0.128,远远低于其它细胞器0.913。蛋白质二级结构预测表明,该蛋白由36.48%的α-螺旋(Alpha helix)、16.67%的延伸链(Extended strand)、7.23% β-转角(Beta turn)和39.62% 的无规则卷曲(Random coil)组成(图3)。SWISS-MODEL三维建模表明,该蛋白含有一个螺旋-环-螺旋结构,具有bHLH家族典型的结构域(图4)。
|
图 3 苦荞转录因子FtbHLH3 的二级结构预测
注: 竖线从短至长依次表示无规则卷曲, β- 转角, 延伸链和α 螺旋
Figure 3 Secondary structure prediction for transcription factor FtbHLH3 from F. tataricum
Note: The vertical lines from short to long mean random coil, beta turn, extended strand and alpha helix
|
.png)
|
图 4 苦荞FtbHLH3 蛋白三维空间结构预测
Figure 4 Tertiary structure prediction of FtbHLH3 proteins from F. tataricum
|
.png)
1.3编码蛋白FtbHLH3的结构分析
Protein Blast分析表明,FtbHLH3序列其它植物的bHLH蛋白同源性在50%~60%。DNAMAN进行的多重比对和NCBI在线工具CD Search分析结果显示(图5; 图6),FtbHLH3与其他植物bHLH转录因子在N端和C端都存在着明显的保守序列,其N端包含有一个可以特异性结合靶基因启动子区域中顺式作用元件E-box元件(5’-CANNTG-3’)的保守域。下游C端含有一个由2个含有疏水性氨基酸的偶极性α螺旋组成HLH结构域。
|
图 5 苦荞转录因子FtbHLH3 与其他植物bHLH 多重序列比对
注: 多重序列比对植物的蛋白序列分别为: 山葡萄: VabHLH (AFH68208); 葡萄: VvbHLH (AFR78197); 莲: NnbHLH93 (XP_010-254652); 苜蓿: MtbHLH93 (XP_003612938)
Figure 5 Multiple sequence alignment between tartary buckwheat FtbHLH3 and other plant bHLHs
Note: The amino acid sequences used to multiple sequence alignment: Vitis amurensis: VabHLH (AFH68208); Vitis vinifera: VvbHLH (AFR78197); Nelumbo nucifera: NnbHLH93 (XP_010254652); Medicago truncatula: MtbHLH93 (XP_003612938)
|
.png)
|
图 6 FtbHLH3 保守结构域分析
Figure 6 FtbHLH3 conserved domains search
|
.png)
1.4编码蛋白FtbHLH3的系统进化分析
将编码蛋白FtbHLH3序列与Blast结果中相似性较高的其他植物已知bHLH序列经采用MEGA 5.0邻接法构建系统发育进化树(图7)。结果表明,该进化树从功能上分为两个大簇,紫苏PfMYC-RP、金鱼草AmDEL、拟南芥AtGL3、苦荞FtbHLH1、葡萄VvMYC1共聚于ClusterⅠ,该簇均是参与植物花青素代谢调控。而苦荞FtbHLH3跟参与植物在非生物逆境胁迫条件的应答相关的枳PtrbHLH、拟南芥AtICE1/AtICE1和亚麻CsICE1转录因子归于ClusterⅡ,推测苦荞FtbHLH3具有相似功能。
|
图 7 苦荞FtbHLH3 与其他植物bHLH 蛋白序列的系统进化树
Figure 7 Phylogenetic tree based on amino acids equences of FtbHLH3 and other plants
|
.png)
1.5非生物逆境胁迫下苦荞FtbHLH3的表达分析
1.5.1 UV-B对苦荞FtbHLH3表达的影响
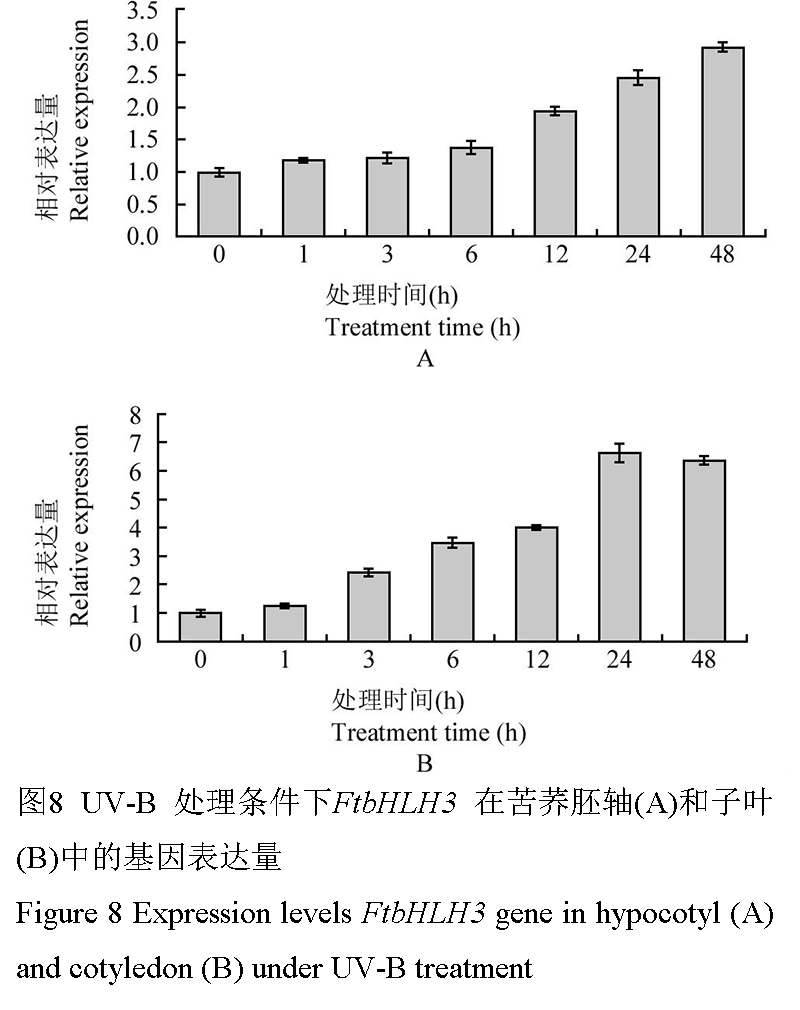
荧光定量PCR对FtbHLH3在UV-B处理后芽期苦荞胚轴和子叶中的相对表达量分析表明(图8),处理后6 h内,胚轴中FtbHLH3的表达量缓慢增加,至处理12 h后明显上升至相对表达量1.95,48 h时达到最大值2.93;而子叶中FtbHLH3的表达量变化则更为明显,1~24 h内该基因的表达量持续显著升高,24 h时达到最大值6.64,之后趋于稳定。
|
图 8 UV-B 处理条件下FtbHLH3 在苦荞胚轴(A)和子叶(B)中的基因表达量
Figure 8 Expression levels FtbHLH3 gene in hypocotyl (A) and cotyledon (B) under UV-B treatment
|
1.5.2 冷胁迫对苦荞FtbHLH3表达的影响
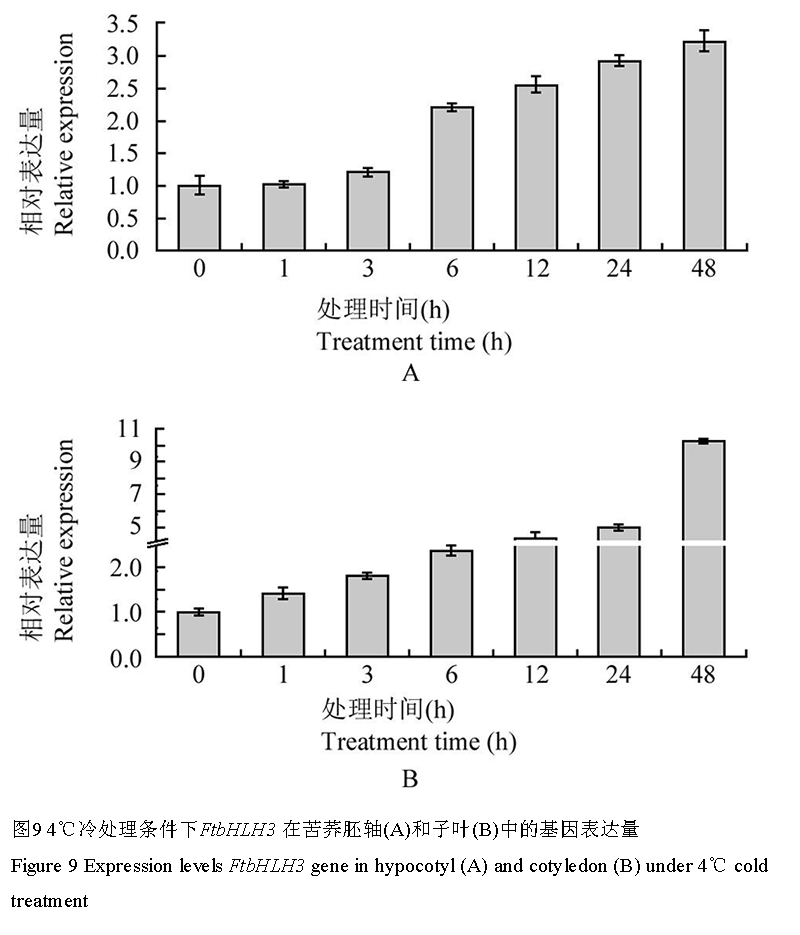
4℃冷胁迫处理后,芽期苦荞胚轴和子叶中FtbHLH3表达量分析显示(图9),胚轴FtbHLH3的表达量在处理3 h内缓慢上升,至6 h时明显上升至相对表达量2.21,48 h时达到最大值3.22;而在子叶中FtbHLH3的表达量则表现出对冷胁迫更强更迅速的应答,3 h即上调至1.81,随后持续大幅度升高,48 h时表达量达最大值10.27。
|
图 9 4℃冷处理条件下FtbHLH3 在苦荞胚轴(A)和子叶(B)中的基因表达量
Figure 9 Expression levels FtbHLH3 gene in hypocotyl (A) and cotyledon (B) under 4℃ cold treatment
|
2讨论
bHLH转录因子作为植物中庞大的转录因子家族之一,在植物代谢和发育的各个方面起着十分重要的调控作用。本研究根据苦荞花期转录组数据,克隆得到苦荞转录因子基因FtbHLH3的cDNA序列。该基因编码蛋白含有bHLH类转录因子特有的保守域,含有一个螺旋-环-螺旋结构,结构上属于bHLH转录因子家族。系统进化树分析显示,FtbHLH3与参与植物抗逆调控的AtIC1 (NP_189309) (Grant et al., 2000)、PtrbHLH (AFY17139) (Huang et al., 2013)等蛋白共聚一簇,而FtbHLH3基因在芽期苦荞胚轴和子叶中对UV-B和冷胁迫的表达变化也进一步说明该蛋白可能参与了苦荞非生物胁迫的调控。
已有研究证明,植物对外界非生物胁迫的应答是一个复杂的过程,植物通过转录因子参与调解的对非生物逆境胁迫的应答受两大类信号通路调控:ABA介导途径与ABA非依赖途径(Shinozaki and Yamaguchi-Shinozaki, 1997; Yamaguchi-Shinozaki and Shinozaki, 1993)。Kiribuchi等(Kiribuchi et al., 2005)报道外源JA诱导水稻bHLH类转录因子RERJ1基因在营养生长期的叶片、叶鞘和根中表达,而低温伤害和干旱胁迫下RERJ1基因在叶片中上调表达。另外,在拟南芥中发现bHLH蛋白rd22BP1能够参与ABA信号途径的脱水胁迫反应(Abe et al., 1997)。本研究也显示了类似结果,即在UV-B和4℃冷两种胁迫处理下苦荞FtbHLH3基因在胚轴和子叶中的表达量均呈持续增长的趋势。此外,FtbHLH3基因在子叶的表达量随着胁迫的处理变化程度则更为显著,这可能是由于植物幼苗在UV-B与冷胁迫下,子叶相比胚轴来说总是以更大的面积受到逆境刺激(Kim et al., 2007)。
苦荞主要分布在我国西南山区,气候干旱、昼夜温差大、紫外线强,而苦荞表现出了良好的抗逆性。本研究克隆得到一条与苦荞抗逆相关的FtbHLH3基因,分析了其在UV-B胁迫和4℃冷胁迫下的响应,为进一步探究苦荞的抗逆机制奠定了基础。
3材料与方法
3.1材料及主要试剂
在西昌学院购买的“西荞2号”作为实验品种,培养条件为光照16 h,黑暗8 h,温度(22± 2)℃,相对湿度70%左右的光照培养箱。植物RNAout试剂盒(天泽基因),Prime Script RT reagent Kit With Gdna Eraser (Perfect Real Time, 宝生物),YBR Premix Ex TaqTM II (TliRNaseH Plus, 宝生物)。其他化学药品均为进口或国产分析纯试剂。
3.2苦荞RNA 提取及FtbHLH3 cDNA 序列的克隆
参照天泽基因公司“植物RNAout 试剂盒”的使用方法来提取花期苦荞子叶的总RNA。再按照RevertAidTM First Strand cDNA Synthesis Kit试剂盒的说明,反转录获得第一链cDNA。根据苦荞花期转录组数据,设计一对特异引物FtbHLH3f和FtHLH3r,以苦荞cDNA第一条链为模板,PCR扩增FtHLH3 cDNA序列,PCR反应条件:98°C 10 sec;98°C 10 sec,59°C 10 sec, 72°C 15 sec,30个循环;72°C 10 min。PCR产物经T载体克隆,筛选阳性菌落,送擎科基因有限公司测序。
3.3 FtbHLH3基因核酸与蛋白质序列分析
通过NCBI在线工具Blast进行FtbHLH3核酸序列分析;利用Prot Param 软件对氨基酸序列的多个物理和化学参数(分子量,等电点,吸光系数等)进行计算。氨基酸亲疏水性特征分析采用Prot Scale程序;蛋白信号肽预测用Signal P程序;利用Tmpred软件预测蛋白跨膜区域;亚细胞定位采用Target P程序进行预测。使用GOR软件进行蛋白质二级结构的预测;采用SWISS-MODEL预测蛋白的三级结构;使用DNAMAN进行多序列比对;利用MEGA5.0 将比对结果采用邻接法(neighbor-joining, NJ)构建系统进化树。
3.4芽期苦荞的胁迫处理
UV-B 处理: 使用10W UV-B (308 nm) LED灯照射芽期苦荞,光照距离 30cm;4℃冷处理: 将芽期苦荞转移至培养条件为光照16 h,黑暗8h,温度4℃,相对湿度60%的光照培养箱进行冷胁迫。以上处理均于处理前0 h、处理后1、3、6、12、24 和 48 h分别取供试样品的胚轴和子叶,液氮冷冻后置于-80℃备用。
3.5苦荞FtbHLH3基因表达量的检测
提取各处理样品的总RNA,通过琼脂糖凝胶电泳检测其完整性,再以RNA为模板反转录合成cDNA。设计一对实时定量PCR特异引物FtbHLH3qf和FtbHLH3qr,以苦荞组蛋白H3基因为内参基因,引物为H3qf和H3qr (表1)。PCR反应体系(15 μL)为:SYBRII 7.5 μL,模板1 μL,上下游引物各0.5 μL,10.5 μL ddH20,反应参数: 95°C 3 min;95°C 10 s, 60°C 30 s, 72°C 30 s, 40个循环;在温度为65~95℃之间绘制溶解曲线,每个反应重复3次后求其平均值,再使用2-ΔΔCt算法来计算基因的相对表达量。
|
表 1 引物序列
Table 1 Primer sequences
|
.png)
作者贡献
姚攀锋负责苦荞FtbHLH3基因的克隆、数据分析及撰写,并参与整个实验过程;赵学荣负责苦荞非生物胁迫处理;李茂菲负责荧光定量PCR;王安虎提供试验材料;吴琦负责论文设计、指导和论文修改;全体作者均已阅读并同意发表。
致谢
本研究由四川省国际合作项目(2015HH0047)资助。
Abe H., Yamaguchi-Shinozaki K., Urao T., Iwasaki T., Hosokawa D., & Shinozaki K.,1997, Role of Arabidopsis MYC and MYB homologs in drought-and abscisic acid-regulated gene expression, The Plant Cell, 9(10), 1859-1868.
Bai Y.C., Li C.L., Zhang J.W., Li S.J., Luo X.P., Yao H.P., Chen H., Zhao H.X., Park S.U., and Wu Q., 2014, Characterization of two tartary buckwheat R2R3-MYB transcription factors and their regulation of proanthocyanidin biosynthesis, Physiologia Plantarum, 152(3): 431-440.
Bonafaccia G., Marocchini M., & Kreft I., 2003, Composition and technological properties of the flour and bran from common and tartary buckwheat, Food chemistry, 80(1), 9-15.
Ding W., Yu Z., Tong Y., Huang W., Chen H., & Wu P., 2009, A transcription factor with a bHLH domain regulates root hair development in rice, Cell research, 19(11): 1309-1311.
Grant D., Cregan P., Shoemaker R.C., 2000, Genome organization in dicots: genome duplication in Arabidopsis and synteny between soybean and Arabidopsis, Proceedings of the National Academy of Sciences, 97(8), 4168-4173.
Huang Y.J., Deng R.Y., Gao F., Luo X.P., and Wu Q., 2015, Cloning and Expression Analysis of Transcription Factor Gene FtMYB21 from Tartary Buckwheat under Abiotic Stress, Jiyinzuxue Yu Yingyong Shengwuxue (Genomics and Applied Biology), 34(9), 1939-1945(黄云吉, 邓仁榆, 高飞, 雒晓鹏, & 吴琦. (2015), 苦荞转录因子基因 FtMYB21 的克隆及其非生物胁迫下的表达分析. 基因组学与应用生物学, 34(9), 1939-1945)
Kim S.J., Maeda T., Takigawa S., Matsuura-Endo C., Yamauchi H., and Mukasa Y., 2007, Identification of anthocyanins in the sprouts of buckwheat, J, Agric.Food Chem, 55(15): 6314-6318.
Kim S.L., Son Y.K., Hwang J.J., Kim S.K., Hur H.S., and Park C.H., 2001, Development and utilization of buckwheat sprouts as functional vegetables, Fagopyrum, 18, 49-54.
Kiribuchi K., Jikumaru Y., Kaku H., Minami E., Hasegawa M., Kodama O., Seto H., Okadad K., Nojiri H., Yamane H., 2005, Involvement of the basic helix-loop-helix transcription factor RERJ1 in wounding and drought stress responses in rice plants, Bioscience, biotechnology, and biochemistry, 69(5), 1042-1044.
Li T., Sun J.K., and Liu J.T., 2015, Role of different transcription factor families in the regulatory networks of drought and salinity tolerance in plants, Shengming Kexue (Chinese Bulletin of Life Sciences), 27(2): 217-227(李田, 孙景宽, 刘京涛, 2015, 植物转录因子家族在耐盐抗旱调控网络中的作用, 生命科学, 27(2): 217-227)
Matus J.T., Poupin M.J., Canon P., Bordeu E., Alcalde J.A., & Arce-Johnson P., 2010, Isolation of WDR and bHLH genes related to flavonoid synthesis in grapevine (Vitis vinifera L.). Plant molecular biology, 72(6), 607-620.
Salvatierra A., Pimentel P., Moya-León M.A., & Herrera R., 2013, Increased accumulation of anthocyanins in Fragaria chiloensis fruits by transient suppression of FcMYB1 gene, Phytochemistry, 90, 25-36.
Sha W., Suo L., Zhang M.J., Zhang C.L., and Zhou B., 2015, Cloning and expression analysis of MYB transcription factor gene RcMYB from Racomitrium canescens, Jiyinzuxue Yu Yingyong Shengwuxue (Genomics and Applied Biology),34(4): 792-796(沙伟, 索荔, 张梅娟, 张春蕾, 周伯, 2015, 砂藓MYB转录因子基因RcMYB的克隆及表达分析, 基因组学与应用生物学, 34(4): 792-796)
Shinozaki K., and Yamaguchi-Shinozaki K., 1997, Gene expression and signal transduction in water-stress response, Plant Physiology, 115(2): 327.
Yamaguchi-Shinozaki K., and Shinozaki K., 1993, The plant hormone abscisic acid mediates the drought-induced expression but not the seed-specific expression of rd22, a gene responsive to dehydration stress in Arabidopsis thaliana, Molecular and General Genetics MGG, 238(1-2): 17-25.
Zhou H., Zhu Q., Yang Y.F., Liu H.W., Yu F.X., Qiu D.Y., 2015, Cloning and Sequence Analysis of bHLH Gene from Taxus chinensis var. mairei, Bulletin of Botanical Research, 35(1), 52-59(周华, 朱祺, 杨艳芳, 刘洪伟, 余发新, & 邱德有. (2015). 南方红豆杉 bHLH 基因克隆与序列分析. 植物研究, 35(1), 52-59)
Zhou M.L., Hou H.L., Zhu X.M., Shao J.R., Wu Y.M., & Tang Y.X., 2011, Soybean transcription factor GmMYBZ2 represses catharanthine biosynthesis in hairy roots of Catharanthus roseus, Applied microbiology and biotechnology, 91(4), 1095-1105.
Zhou M.L., Wang C.L., Qi L.P., Yang X.B., Sun Z.M., Tang Y., Tang Y.X., Shao J.R., and Wu Y.M., 2015, Ectopic expression of Fagopyrum tataricum FtMYB12 improves cold tolerance in Arabidopsis thaliana, Journal of Plant Growth Regulation, 34(2): 362-371.