研究报告/Research Report
黑果枸杞和宁夏枸杞FT (Flowering Locus T)基因的克隆及表达分析 
2中国科学院大学, 北京, 100039;
3中国科学院华南植物园, 中国科学院植物资源保护与利用重点实验室, 广东省应用植物学重点实验室, 广州, 510650
 作者
作者  通讯作者
通讯作者
植物药与药理学杂志, 2014 年, 第 3 卷, 第 4 篇
收稿日期: 2014年03月05日 接受日期: 2014年03月15日 发表日期: 2014年03月25日
引用格式(中文):
薛定磊等, 2014, 黑果枸杞和宁夏枸杞FT (Flowering Locus T)基因的克隆及表达分析, 基因组学与应用生物学, 34(3): 565-570 (doi: 10.13417/j.gab.034.000565)
引用格式(英文):
Xue et al., 2014, Molecular Cloning and Expression Analysis of Flowering Locus T in Lycium ruthenicum Murr. and L. barbarum L, Genomics and Applied Biology, 34(3): 565-570 (doi: 10.13417/j.gab.034.000565)
黑果枸杞(Lycium ruthenicum Murr.)和宁夏枸杞(Lycium barbarum L.)系茄科枸杞属植物,具有非常重要的药用价值。本研究通过挖掘黑果枸杞和宁夏枸杞的转录组数据克隆获得其开花关键基因FT (Flowering Locus T),分别命名为LrFT和LbFT,通过生物信息学方法分析两个蛋白质的氨基酸序列、功能域及其系统发育关系等;并通过实时定量PCR分析其在两种枸杞不同组织中的表达模式。研究结果表明:LrFT和LbFT均编码一个由173个氨基酸组成的蛋白质。蛋白序列分析表明:LrFT和LbFT序列相似度达98%,具有保守的PEBP (phosphatidylethanolamine-binding protein)结构域。系统进化分析表明:LrFT和LbFT与茄科植物番茄、烟草和马铃薯的FT亲缘关系较近。基因表达结果表明:LrFT和LbFT主要在叶片中表达,且在两种枸杞的叶片中表达量有一定的差异。本研究为今后黑果枸杞和宁夏枸杞开花相关功能基因的验证及分子育种奠定了一定的基础。
黑果枸杞(Lycium ruthenicum Murr.)和宁夏枸杞(Lycium barbarum L.)系茄科(Solanaceae)枸杞属浆果类植物,是我国传统的名贵中药材。黑果枸杞,藏药称为“旁玛”,因果实成熟后呈黑色而得名(甘青梅等, 1997)。其果实味甘、性平、清心热,藏药用于治疗心脏病、月经不调、停经及心热病等,被收录于《晶珠本草》和《四部医典》等药典(甘青梅, 2001)。汪建红等(2009)研究发现黑果枸杞的果实多糖具有降血糖、抗疲劳等功效。此外,黑果枸杞抗逆性极强,是我国西北地区用于改善土地荒漠化的优良植物资源(陈海魁等, 2008)。宁夏枸杞也是一种重要的药用植物,是《中国药典》中地骨皮和枸杞子的基源植物(国家药典委员会, 2010)。早在《神农本草经》、《本草纲目》等古代医学名著中就有枸杞可入药的记载。现代医学研究表明,宁夏枸杞具有抗衰老(Li et al., 2007)、抗肿瘤(Chao et al., 2006)、抗辐射(Qian et al., 2004)、养肝明目(Hsu et al., 1999)、强筋健骨(Yu et al., 2005)、增强免疫力(Lin et al., 2008)等功效。
目前,我国对黑果枸杞的研究主要集中在其组织培养(浩仁塔本等, 2005)及活性成分原花青素(Zheng et al., 2011)、多糖(Peng et al., 2012)、甜菜碱(刘增根等, 2012)、类黄酮(李淑珍等, 2008)的药效药理等方面;而对宁夏枸杞的研究则主要集中在其栽培、生产加工、药物活性成分分离及鉴定(Lim, 2013)等方向。但是,对于黑果枸杞和宁夏枸杞开花的分子机理等方面的研究却没有报道。由于黑果枸杞和宁夏枸杞的童期较长,生长周期至少一年,不利于在实验室条件下进行以其果实为研究对象的次生代谢调控等研究。
关于植物开花机理的研究,最早可追溯到1865年Sachs首次提出成花物质的概念,并分析指出该物质能由植物的叶片运输到叶芽导致植物开花(张婧, 2012)。为了证明这一结论,人们先后提出了开花物质假说、营养物质转移假说及多因子控制模型等解释植物开花的机理(郭春晓等, 2009)。其中,营养物质假说(Sachs and Hackete, 1972)认为适当的诱导处理可以导致植物叶片大量的同化产物在顶端分生组织积累,从而导致其开花;多因子控制模型(Bernier, 1988)则是在营养物质假说的基础上提出的,它认为营养物质的积累仅仅只是诱导植物开花的一个方面,除此之外还有其它诸如激素等都会在植物开花过程中起作用。近期,人们发现了控制植物开花的关键基因FT (FLOWERING LOCUS T)。随后的研究指出植物开花中真正起作用的是FT蛋白,而不是FT mRNA (Corbesier et al., 2007)。Mitsutomo等(2005)进一步研究发现拟南芥在接受适当的光周期诱导后,叶片产生的FT mRNA经叶片维管组织运输到茎顶端分生组织,翻译后生成的FT蛋白与bZIP转录因子FLOWERING LOCUS D (FD)互相作用,激活花分生组织基因APETALA1 (AP1)的表达,从而促进成花转换、启动花的发育过程。
本文通过挖掘两种枸杞的转录组数据,克隆了控制黑果枸杞和宁夏枸杞开花的关键基因FT,分别命名为LrFT和LbFT,并对其进行序列分析和表达分析,为两种枸杞种质资源改良及次生代谢调控等方面的研究提供一定的理论基础,对后期进一步研究其药用价值和种质资源的选育具有显著意义。
1结果与分析
1.1 LrFT和LbFT及其蛋白序列分析
利用拟南芥FT基因序列检索黑果枸杞和宁夏枸杞EST数据库,最终获得FT基因各一个拷贝,并命名为LrFT和LbFT;且序列分析表明,这两个FT基因都编码全长基因。根据FT基因已知序列设计全长引物,分别以黑果枸杞和宁夏枸杞幼嫩叶片cDNA为模板,经PCR克隆、测序验证结果表明LrFT和LbFT编码区序列长度均为522 bp,各有一个完整的阅读框,编码173个氨基酸。ExPASy在线工具(http://web.expasy.org/compute_pi/)预测结果表明: LrFT和LbFT蛋白的等电点分别为9.10和9.23,分子量分别为18.76 KD和18.87 KD。氨基酸序列比对结果表明LrFT与LbFT序列一致性达98%。LrFT和LbFT的氨基酸序列与同属茄科的马铃薯、烟草的FT序列也有较高的一致性(图1),表明LrFT和LbFT克隆的正确性。
.png) 图1 LrFT, LbFT与其它物种蛋白序列比对 |
1.2 LrFT和LbFT系统进化分析
从NCBI数据库中搜索获得葡萄(Vitis vinifera)、苹果(Malus pumila)、番茄(Solanum lycopersicum)等8个物种的FT蛋白序列与LrFT和LbFT蛋白序列一起构建NJ进化树(图2)。系统进化分析结果表明,LrFT和LbFT同属一支,亲缘关系最近。此外,从系统进化树可以看出,LrFT和LbFT与同属茄科的烟草和马铃薯的FT蛋白(Nicotiana_tomentosiformis, AGJ83936; Solanum_tuberosum, AGH33864)聚为一类,这与它们同属茄科相一致。
.png) 图2 LrFT和LbFT与其它物种FT蛋白系统进化分析 |
1.3 LrFT和LbFT蛋白同源建模
利用Swiss-Model的Alligenment Model在线工具(http://www.swissmodel.expasy.org/interactive#alignment)构建了LrFT和LbFT的三维模型。蛋白三维模型表明:LrFT和LbFT的三维结构无明显差异,表现出高度的相似性(图3)。
.png) 图3 LrFT(A)和LbFT(B)蛋白三维结构比较 |
1.4 LrFT和LbFT蛋白功能域分析
利用DNAMAN 6软件,将LrFT和LbFT与AtFT (拟南芥)、CiFT (柑橘)、VvFT (葡萄)、StFT (马铃薯)、NtFT (烟草)进行氨基酸序列比对,并通过NCBI Conserved Domains在线数据库(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)对LrFT和LbFT保守区域进行预测,如图1和图4所示:LrFT和LbFT都包含一个保守的PEBP结构域。该结构域由5个方向平行的肽链形成一个大的中心β-折叠片和连接在两侧的1个较小的β-折叠片和1个C端α-螺旋组成;其中在中心β-折叠片的一端有1个独特且高度保守的阴离子结合位点该位点负责识别磷酸化的残基,是该结构域的主要功能域(Hanzawa et al., 2005; Ahn et al., 2006)。
.png) 图4 LrFT(A)和LbFT(B)保守功能域的预测分析 |
1.5 FT基因在不同器官中的表达
基因表达结果表明,FT基因主要在黑果枸杞和宁夏枸杞的叶片中表达,而在根、茎、花等器官中表达量较低(图5)。目前关于植物开花的研究指出,植物叶片接受光周期诱导后能够激活FT基因的表达,其编码的蛋白能够通过叶片韧皮部,由叶片运输到茎端分生组织,与bZIP转录因子FD蛋白结合,上调SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1)基因和花分生组织特性基因AP1的表达,促进花分生组织的形成而开花(Mitsutomo et al., 2005; Wigge et al., 2005)。
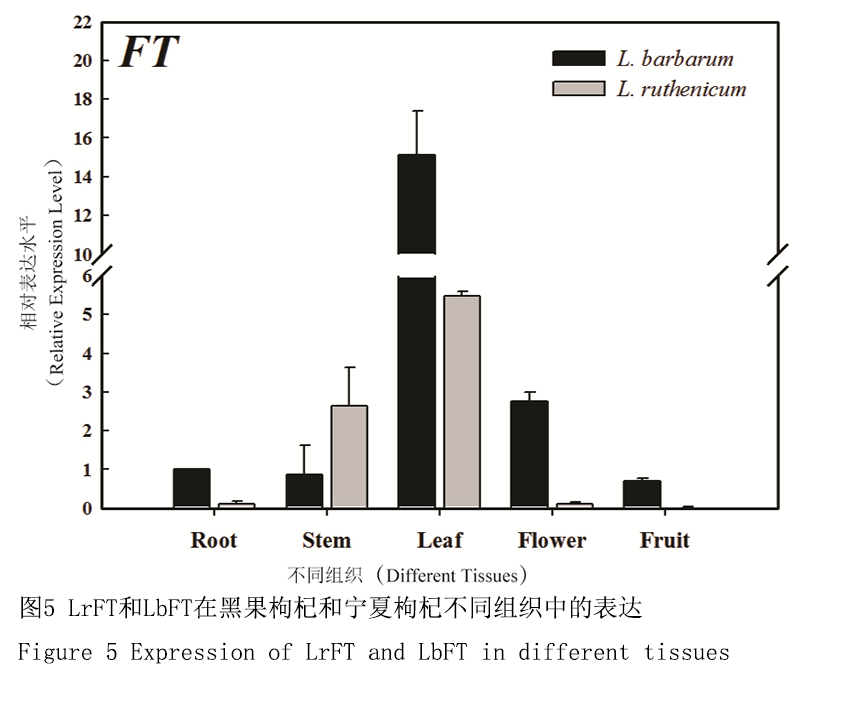 图5 LrFT和LbFT在黑果枸杞和宁夏枸杞不同组织中的表达 |
2讨论
高等植物由营养生长向生殖生长转换的过程我们称之为成花转变,主要有开花诱导、花的发端和花器官发育三个阶段;是一个由内部遗传因子和外部环境因子相互协调控制的多基因调控过程(Bernier, 1988)。这个过程决定了植物的开花时间,而植物的开花时间不仅决定了植物在某一特定生态区域的适应性,还间接的影响到人类的生活。因此,通过研究植物开花的分子机制可以实现对植物开花时间的有效调控,从而解决实际生产中遇到的诸如育种、制种中存在的花期不遇等问题,为优良种质资源的性状改良提供可靠的理论依据。
FT基因是花发育途径的汇集点,它能够整合来自光周期途径(photoperiodic pathway)、春化途径(vernalization pathway)和自主途径(autonomous pathway)等不同花发育途径的信号(Corbesier and Coupland, 2006),在转录因子FD (flowering locus D)作用下,上调SOC1基因(Moon et al., 2003)和花分生组织特异基因APETALA 1 (AP1) (Liljegren et al., 1999)的表达,促进花原基的形成,在植物花发育过程中具有重要作用。本研究通过转录组数据设计引物分别克隆获得控制黑果枸杞和宁夏枸杞开花的关键因子FT基因,分别命名为LrFT和LbFT。通过氨基酸序列比对发现LrFT与LbFT序列相似度高达98%,并且它们和马铃薯、烟草的氨基酸序列相似度也非常高,这与它们同属茄科相一致。蛋白质结构域及功能域分析表明LrFT和LbFT具有共同的PEBP结构域,该结构域在植物FT/TFL1基因家族是高度保守的,因其能在体外结合磷脂酰乙醇胺而得名。基因表达分析表明,LrFT和LbFT两个基因主要在两种枸杞的嫩叶中表达,且在宁夏枸杞嫩叶中的表达量要高于黑果枸杞。此外,在黑果枸杞和宁夏枸杞的根、茎、花和果实中,LrFT和LbFT也有表达,但表达量都相对较低,这与拟南芥等多数模式植物的FT同源基因表达模式基本一致,同时与FT基因编码的蛋白可以通过叶片韧皮部运输到茎顶端分生组织和FD蛋白结合进而诱导植物开花的研究相一致(Mitsutomo et al., 2005; Wigge et al., 2005; Corbesier et al., 2007)。
由于黑果枸杞和宁夏枸杞的药用部位主要是果实;而黑果枸杞和宁夏枸杞均系多年生灌木,花期5~10月,正常植株从幼苗到开花结果至少需要一年的生长周期;因此,不利于枸杞种质资源的改良和经济效益的提高。LrFT、LbFT基因全长的克隆获得,为今后黑果枸杞和宁夏枸杞早花株系的培育、促进果实品质性状的研究奠定了基础。
3材料与方法
3.1材料
黑果枸杞和宁夏枸杞样品均来自宁夏中宁县实验基地,分别取同一棵植株上的根、茎、嫩叶、花瓣和果实若干,经液氮冷冻后置于-80℃低温冷藏。
3.2 LrFT和LbFT基因克隆
Trizol法依次提取两种枸杞的根、茎、嫩叶、花瓣和果实的总RNA,反转录法合成cDNA,稀释10倍作为模板。根据实验室提供的两种枸杞的转录组数据经序列拼接注释,从中分别发掘出其对应的FT基因的转录本,设计引物,正向:5'-AACATATTTCCCTCTAGCTAATGGC-3';反向:5'-GTTACTTCTTAGCGCTTACGATTTG-3'。PCR反应体系为25 μL:10×La Buffer 2.5 μL,dNTP 4.0 μL,上下游引物(10 μmol/L)各1.0 μL,cDNA模板0.5 μL,TaKaRa La Taq (2.5 U/μL) 0.25 μL,灭菌双蒸水15.75 μL。PCR反应程序如下:95℃ 5 min;94℃ 30 s;55℃ 30 s;72℃ 1 min,35个循环。PCR产物经凝胶电泳分析并回收。将回收产物克隆到pMD19-T载体(购自TaKaRa公司)上并转化大肠杆菌DH5α超级感受态细胞,涂板,37℃隔夜培养。次日挑取单菌落于500μL LB液体培养基(含AMP 0.05 mg/L),200 r,37℃摇床培养6 h,菌液PCR检测并将获得的阳性菌株送样至上海生工生物公司测序验证。
3.3 LrFT和LbFT序列及编码蛋白的分析
ExPASy在线工具(http://web.expasy.org/compute_pi/)对LrFT和LbFT的氨基酸理化性质进行预测分析;DNAMAN 6软件比对分析LrFT和LbFT的氨基酸序列;NCBI Conserved Domains在线数据库(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析LrFT和LbFT蛋白保守功能域;Swiss-Model中的alignment model在线工具(http://swissmodel.expasy.org/interactive)对LrFT和LbFT进行蛋白同源建模;MEGA 5.1软件构建LrFT和LbFT的NJ系统进化树,选用默认参数。选取以下物种的FT蛋白进行系统发育树分析,葡萄(Vitis vinifera, VvFT, DQ871590; grapevine, VvFT, XP_002270408);苹果(Malus x domestica, MdFT, AB161112);柑橘(Citrus reticulata Blanco, CiFT, AB027456; CiFT2, AB301934.1);拟南芥(Arabidopis thaliana, AtFT, AB027504);杨树(Populus simonii Carriere, PnFT2a, BAD01561; PtFT2, AFU08240);番茄(Solanum lycopersicum, FT, XP_004239845);烟草(Nicotiana tomentosiformis, FT, AGJ83936);马铃薯(Solanum tuberosum, FT, AGH33864)。
3.4 FT基因的表达分析
利用prime 5.0设计实时定量PCR引物,正向引物:5'-TGGACATGTTCGCCTAGT-3',反向引物:5'-TTCACTTGGACTGGGAGCAT-3';其中,内参actin引物由沈笑飞等(2014)提供。PCR总反应体系为20 μL:正反向引物(10 μmol/L)各0.8 μL,灭菌双蒸水6.4 μL,SYBR® Premix Ex Taq™ II (Tli RnaseH Plus) 10 μL,模板cDNA 2 μL。实时定量PCR参数:95℃ 30 s;95℃ 5 s,60℃ 20 s,72℃ 20 s,40个循环。采用Livak和Schmittgen (2001)报道的双△法计算相对表达量,每个样品重复3次,以内参actin基因计算FT基因的相对表达量。
作者贡献
通讯作者负责指导实验及修改论文;薛定磊负责实施实验、分析数据及撰写文章;曾少华负责采集样品。
致谢
感谢国家自然科学基金(31470391)和中国科学院科研装备研制项目(YZ201227)对本课题相关研究的资助。
参考文献
Ahn J.H., Miller D., Winter V.J., Banfield M.J., Lee J.H., Yoo S.Y., Henz S.R., Brady R.L., and Weigel D., 2006, A divergent external loop confers antagonistic activity on floral regulators FT and TFL1, The EMBO Journal, 25(3): 605-614
Bernier G., 1988, The control of floral evocation and morphogenesis, Annu. Rev. Plant Physiol. Plant Mol. Biol., 39: 175-219
Chao J.C.J., Chiang S.W., Wang C.C., Tsai Y.H., and Wu M.S., 2006, Hot water-extracted Lycium barbarum and Rehmannia glutinosa inhibit proliferation and induce apoptosis of hepatocellular carcinoma cells, World Journal of Gastroenterology, 12(28): 4478-4484
Chen H.K., Pu L.K., Cao J.M., and Ren X., 2008, Current research state and exploitation of Lycium ruthenicum Murr, Heilongjiang Nongye Kexue (Heilongjiang Agricultural Sciences), (5): 155-157 (陈海魁, 蒲淩奎, 曹君迈, 任贤, 2008, 黑果枸杞的研究现状及其开发利用, 黑龙江农业科学, (5): 155-157)
Corbesier L., and Coupland G., 2006, The quest for florigen: a review of recent progress, J. Exp. Bot., 57(13): 3395-3403
Corbesier L., Vincent C., Jang S., Fornara F., Fan Q., Searle I., Giakountis A., Farrona S., Gissot L., Turnbull C., and Coupland G., 2007, FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis, Science, 316(5827): 1030-1033
Gan Q.M., 2001, Preliminary investigation on Tibetan medicinal herb, Zhongcaoyao (Chinese Traditional and Herbal Drugs), 32(4): 371-373 (甘青梅, 2001, 浅述藏药的研究, 中草药, 32(4): 371-373)
Gan Q.M., Luo G.F., Li P.Y., Zhuoma D.Z., Chen Y.R., and Zuo Z.C., 1997, The study of Tibetan drug Lycium ruthenicum Murr. in development and utilization, Qinghai Keji (Qinghai Science and Technology), 4(1): 17-19 (甘青梅, 骆桂发, 李普衍, 卓玛东珠, 陈岳蓉, 左振常, 1997, 藏药黑果枸杞开发利用的研究, 青海科技, 4(1): 17-19)
Guo C.X., Tian S.B., Zheng C.S., Wang W.L., and Sun X.Z., 2009, FT gene of flowering determination of plants in photoperiod pathway, Jiyinzuxue Yu Yingyong Shengwuxue (Genomics and Applied Biology), 28(3): 613-618 (郭春晓, 田素波, 郑成淑, 王文莉, 孙宪芝, 2009, 光周期途径植物开花决定关键基因FT, 基因组学与应用生物学, 28(3): 613-618)
Hanzawa Y., Money T., and Bradley D., 2005, A single amino acid converts a repressor to an activator of flowering, Proc. Natl. Acad. Sci. USA, 102(21): 7748-7753
Haoren T.B., Zhao Y., Guo Y.S., and Liu P.S., 2005, Tissue culture of Lycium ruthenicum Murr., Zhiwu Shenglixue Tongxun (Plant Physiology Communications), 41(5): 631 (浩仁塔本, 赵颖, 郭永盛, 刘平生, 2005, 黑果枸杞的组织培养, 植物生理学通讯, 41(5): 631)
Hsu H.Y., Yang J.J., Ho Y.H. and Lin C.C., 1999, Difference in the effects of radioprotection between aerial and root parts of Lycium chinense, Journal of Ethnopharmacology, 64(2): 101-108
Li S.Z., Li J., Yang Z.J., and Yuan H., 2008, Technology of extracting and refining flavonoids from Lycium ruthenicum Murr., Shipin Yanjiu Yu Kaifa (Food Research and Development), 29(8): 82-87 (李淑珍, 李进, 杨志江, 原惠, 2008, 黑果枸杞类黄酮的提取和精制工艺研究, 食品研究与开发, 29(8): 82-87)
Li X.M., Ma Y.L., and Liu X.J., 2007, Effect of the Lycium barbarum polysaccharides on age-related oxidative stress in aged mice, J. Ethnopharmacol., 111(3): 504-511
Liljegren S.J., Gustafson-Brown C., Pinyopich A., Ditta G.S., and Yanofsky M.F., 1999, Interactions among APETALA1, LEAFY, and TERMINAL FLOWER1 specify meristem fate, The Plant Cell, 11(6): 1007-1018
Lim T. K., 2013, Lycium barbarum, Edible Medicinal and Non-Medicinal Plants, 6: 240-266
Lin F.Y., Lai Y.K., Yu H.C., Chen N.Y., Chang C.Y., Lo H.C., and Hsu T.H., 2008, Effects of Lycium barbarum extract on production and immunomodulatory activity of the extracellular polysaccharopeptides from submerged fermentation culture of Coriolus versicolor, Food Chemistry, 110(2): 446-453
Liu Z.G., Tao Y.D., Shao Y., and Zhang H.G., 2012, Determination of betaine in Lycium ruthenicum Murr. and Lycium barbarum L., Guangpu Shiyanshi (Chinese Journal of Spectroscopy Laboratory), 29(2): 694-697 (刘增根, 陶燕铎, 邵赟, 张怀刚, 2012, 柴达木枸杞和黑果枸杞中甜菜碱的测定, 光谱实验室, 29(2): 694-697)
Livak K.J., and Schmittgen T.D., 2001, Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method, Methods, 25(4): 402-408
Mitsutomo A., Yasushi K., Sumiko Y., Yasufumi D., Ayako Y., Yoko I., Harutaka I., Michitaka N., Koji G., and Takashi A., 2005, FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex, Science, 309(5737): 1052-1056
Moon J., Suh S.S., Lee H., Choi K.R., Hong C.B., Paek N.C., Kim S.G., and Lee I., 2003, The SOC1 MADS-box gene integrates vernalization and gibberellin signals for flowering in Arabidopsis, The Plant Journal, 35(5): 613-623
Peng Q., Lv X.P., Xu Q.S., Li Y., Huang L.J. and Du Y.G., 2012, Isolation and structural characterization of the polysaccharide LRGP1 from Lycium ruthenicum, Carbohydr Polym, 90(1): 95-101
Qian J.Y., Liu D., and Huang A.G., 2004, The efficiency of flavonoids in polar extracts of Lycium chinense Mill fruits as free radical scavenger, Food Chemistry, 87(2): 283-288
Sachs R.M., and Hackete W.P., 1972, Chemical inhibition of plant height, HortScience, 7: 440-447
Shen X.F., Liu Y.L., Wang Y., and Zeng S.H., 2014, Molecular cloning and expression analysis of flavonol synthase in Lycium ruthenicum Murr. and Lycium barbarum L., Jiyinzuxue Yu Yingyong Shengwuxue (Genomics and Applied Biology), 33(3): 591-597 (沈笑飞, 刘永亮, 王瑛, 曾少华, 2014, 黑果枸杞和宁夏枸杞中黄酮醇合酶基因的克隆及表达分析, 基因组学与应用生物学, 33(3): 591-597)
Wang J.H., Chen X.Q., and Zhang W.J., 2009, Study on hypoglycemic function of polysaccharides from Lycium ruthenicum Murr. fruit and its mechanism, Shipin Kexue (Food Sciences), 30(5): 244-248 (汪建红, 陈晓琴, 张蔚佼, 2009, 黑果枸杞果实多糖降血糖生物功效及其机制研究, 食品科学, 30(5): 244-248)
Wigge P.A., Kim M.C., Jaeger K.E., Busch W., Schmid M., Lohmann J.U., and Weigel D., 2005, Integration of spatial and temporal information during floral induction in Arabidopsis, Science, 309(5737): 1056-1059
Yu M.S., Leung S.K., Lai S.W., Che C.M., Zee S.Y., So K.F., Yuen W.H., and Chang R.C., 2005, Neuroprotective effects of anti-aging oriental medicine Lycium barbarum against beta-amyloid peptide neurotoxicity, Exp. Gerontol., 40(8-9): 716-727
Zhang J., 2012, Research advances on FT gene in higher plant, Xiandai Nongye Keji (Modern Agricultural Sciences and Technology), (3): 31-32 (张婧, 2012, 高等植物开花基因FT研究进展, 现代农业科技, (3): 31-32)
Zheng J., Ding C.X., Wang L.S., Li G.L., Shi J.Y., Li H., Wang H.L., and SuoY.R., 2011, Anthocyanins composition and antioxidant activity of wild Lycium ruthenicum Murr. from Qinghai-Tibet Plateau, Food Chemistry, 126(3): 859-865