人GJB2分子进化与耳聋遗传效应的关联性分析 
2南京医科大学基础医学院, 南京, 210029
 作者
作者  通讯作者
通讯作者
医学遗传学与疾病研究, 2012 年, 第 1 卷, 第 2 篇 doi: 10.5376/mgdr.cn.2012.01.0002
收稿日期: 2012年04月02日 接受日期: 2012年07月23日 发表日期: 2012年08月06日
引用格式(中文):
范燚等, 2012, 人GJB2分子进化与耳聋遗传效应的关联性分析, 医学遗传学与疾病研究(online) Vol.1 No.2 pp.6-14 (doi: 10.5376/mgdr.cn.2012.1.0002)
引用格式(英文):
Fan et al., 2012, Analysis on Association between Molecular Evolution of Human GJB2 Gene and Hereditary Effect with Deafness, Yixue Yichuanxue Yu Jibing Yanjiu (online) Vol.1 No.2 pp.6-14 (doi: 10.5376/mgdr.cn.2012.2.0002)
间隙连接蛋白β2 (GJB2)基因突变与遗传性非综合征性耳聋密切相关,其广泛的突变类型及特异性的热点突变被认为是一种独特的致聋基因。本研究应用生物信息学方法对17个物种的GJB2蛋白进行了系统发育、保守性、跨膜区结构、三维结构和错义突变的分析,并结合已有报道的实验结果进行关联性分析。分析预测获得了166个固定的氨基酸位点,2个非保守区以及2个空间结构保守位点;关联性分析证实发生在保守位点的突变致病性高,非保守区突变的概率致病性小,跨膜区且改变氨基酸性质的突变,可能影响蛋白的空间结构而改变膜通道的通透性。本文为进一步研究GJB2基因突变与聋病的关联性及分子发病机制提供了理论依据,同时,这种研究思路对其它疾病的相关研究具有一定的借鉴价值。
GJB2基因编码的间隙连接蛋白26 (Connexin26, Cx26)是间隙连接蛋白家族中重要的成员,在内耳中是K+和IP3等第二信使分子的通道,对听觉生理和病理具有重要作用。GJB2基因突变与遗传性非综合征型耳聋(nonsyndromic hearing loss, NSHL)有关,是最常见的耳聋基因(Mahdieh et al., 2010; Wu et al., 2011)。在GJB2基因中,己发现100多种突变与耳聋有关(http://davinci.crg.es/deafness/),该基因的突变具有种族和地域特征,如35delG、167delT和235delC分别在高加索人、犹太人和亚洲人群中较为常见。在我国GJB2基因235delC的突变致聋的频率也高于其它亚洲国家(Dai et al., 2007)。由于GJB2基因突变种类多,且仍有新的突变被不断发现,如何科学地预测新生致病突变以及与疾病表型的关联性,是一个值得探索的课题。现今利用生物信息学方法从已有的生物数据库中挖掘有意义的信息,对新发现的突变进行致病性预测和可能的功能分析,此为疾病基因组学研究提供了一个有用的平台。基于此,本文利用生物信息学的方法分析GJB2基因的序列进化和分子特征,并结合突变数据库与相关的研究报道,探讨GJB2基因变异与耳聋遗传效应之间的关系。
1结果与分析
1.1 17个物种及其GJB2蛋白序列信息
本研究选取的17个物种的GJB2蛋白序列信息(表1),包括了15个哺乳纲和2个两栖纲物种,在哺乳纲中包括了灵长目、食肉目、奇蹄目、偶蹄目及啮齿目。选取的17个GJB2蛋白序列中,除了西部爪蛙和非洲爪蛙的GJB2蛋白序列长度为248以外,均为226。
|
|
.png)
1.2 GJB2系统发育分析
从GJB2的系统发育树中可见(图1),哺乳类动物的GJB2与两栖类动物的GJB2亲缘关系较远。此外,在15种哺乳类动物的GJB2、2种两栖类动物的GJB2和哺乳类与两栖类GJB2祖先序列,分别进行的多序列比对结果中,相似度(固定氨基酸残基的频率)分别为83.6% (189/226)、94.0% (233/248)和70.7% (159/225)。
可见,哺乳类动物的祖先序列与两栖类动物的祖先序列相比,相似度较低(图2),与GJB2系统发育树中的结果一致。
|
|
.png)
|
|
.png)
1.3 GJB2保守区分析
在17个物种的GJB2多序列比对中(图2),人类GJB2有65.93% (149/226)的氨基酸残基是固定的,23.89% (54/226)的氨基酸残基是保守的。比较人类的GJB2序列与进化树中各结点处的祖先序列,结果显示,人类GJB2中70.35% (159/226)的氨基酸残基是固定的,20.80% (47/226)的氨基酸残基是保守的。此外,在17个物种序列比对中,其中固定的氨基酸残基在祖先序列中也是一致的。将人类GJB2序列、哺乳动物的祖先序列和两栖类动物的祖先序列进行多序列比对,有73.45% (166/226)的位点显示是固定的。由于基因进化作用,GJB2出现多态性位点从而形成了不同物种,其中部分多态性位点在人类与祖先序列比对中是固定的,因此,将人类GJB2序列中的这些位点视为固定位点可提高分析的精确度。所以,最终得到了166个固定的氨基酸残基位点。
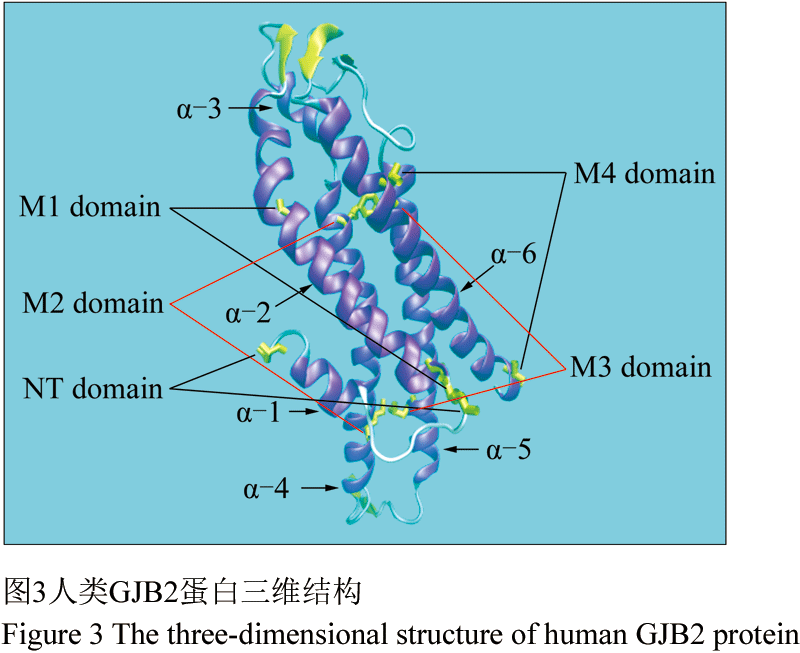
人类GJB2序列的SMART分析(Kelley et al., 1998)显示,GJB2是由5个非跨膜区(2个细胞外环, 1个N末端, 1个C末端和1个细胞内环)和4个跨膜区组成(图3),53↔ 180、60↔ 174及64↔ 169为二硫键结合位点。研究表明,间隙连接蛋白家族成员之间的同源性主要在细胞外的两个结构域,其次是跨膜结构域,细胞质内的结构域差别最大。经比对分析各区域的保守度,结果显示细胞外环区、跨膜区与胞质区的平均保守度分别为94.74%、93.38%和72.78% (表2)。GJB2蛋白细胞外环的高度保守性,推测其可能与间隙连接蛋白其他家族成员在细胞外侧存在相互作用。此外,由于胞质区CT结构域的保守度极低,在本文定义为“1号非保守区”。
|
|
|
|
1.4人类GJB2蛋白三维结构比较
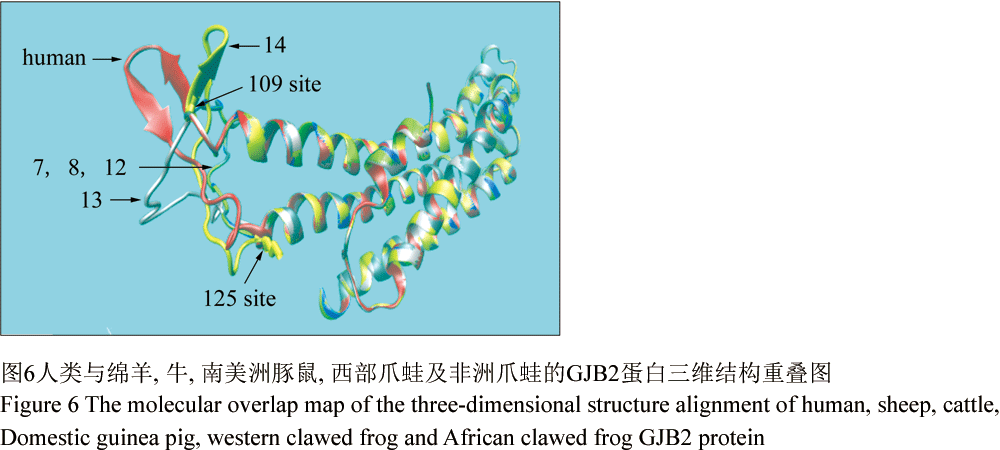
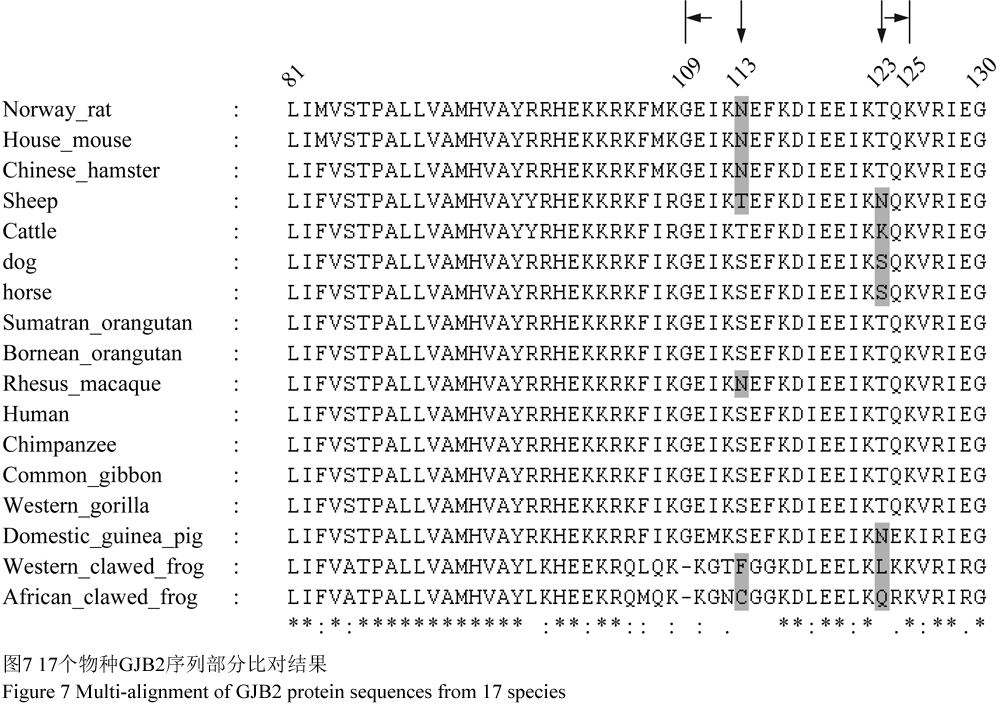
人类GJB2的三维结构包括了6个α螺旋、5个β折叠、10个转角以及7个卷曲(图3)。将人类GJB2三维结构与其余16个物种进行两两比较,得分≥98的序列三维结构与人类的几乎完全重叠,分别是编号为2、3、4、5、6、15、16和17的8个物种(图4)。将比对得分较低的西部爪蛙(score=73)的GJB2三维结构与人类的进行空间比对,发现两者在第109~125氨基酸残基段存在空间结构的差异(图5)。进而,将人类与其余物种的109~125区段分别进行了结构比对,发现物种编号为2、3、4、5、6、9、10、11、15、16及17的三维结构与人类的几乎完全重叠,而编号为7、8、12、13和14的三维结构在109~125残基段与人类存在差异(图6)。分析发现,编号为4、5、6、9、10、11仅在第113位或第123位存在差异,编号7、8、13、14在113和123位同时存在差异,编号12仅在123位有差异,但其与人类亲缘关系较远(图7)。所以,推测当人类GJB2序列的113位和123位同时发生变化时,其序列109~125段对应的空间结构将有可能发生改变。因此,人类GJB2序列第113和123位点可能是“空间结构保守位点”。
|
|
.png)
|
|
.png)
|
|
|
|
由于109~125区段位于99~131非跨膜区域内,其保守度(78.79%)相比其他区域较低(除1号非保守区),在本文将该区域定义为“2号非保守区”。
1.5人类GJB2基因错义突变分析
从CRG (center for genomic regulation)数据库中获得GJB2基因的77种与NSHL相关的致病性错义突变位点信息,其中包括60种常染色体隐性遗传错义突变,8种常染色体显性遗传错义突变和9种功能尚未明确的致病性错义突变(表3)。这些突变共涉及了60个氨基酸位点,其中90% (54/60)为保守性位点。从各区域的分布频率可知(表2),突变位点多数集中在区4 (47.8%),1号非保守区的突变位点分布频率极低(0%),2号非保守区较低(29.4%)。因此,可以推测新的致病性错义突变位点出现在M2结构域的概率较高,出现在CT结构域的概率较低。此外,在上述两个空间结构保守位点均出现了错义突变(S113R和T123N),有研究表明,S113R突变使GJB2不能诱导同型连接通道的形成(Martínez et al., 2009),影响了其膜通道转运功能,生物信息学预测结果也证实该位点的突变将改变GJB2蛋白的三维结构。
|
|
.png)
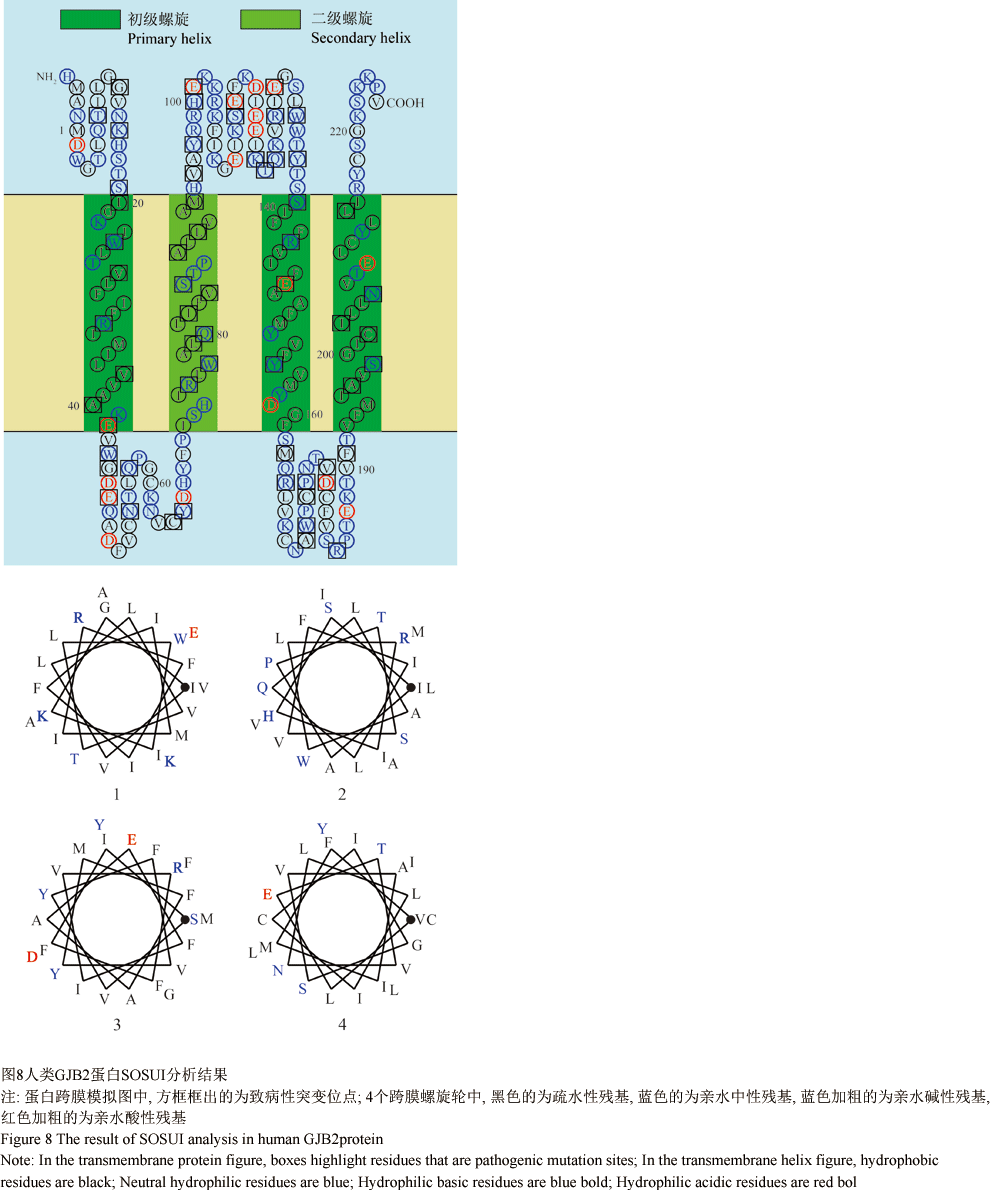
1.6人类GJB2蛋白SOSUI分析
对人类GJB2蛋白进行SOSUI分析,结果显示的跨膜区与SMART分析结果趋于一致。60个突变位点近半数集中在跨膜区(图8),其中23个位于疏水性残基,37个位于亲水性残基。在组成的跨膜区的残基中,28.3% (26/92)为亲水性残基,而分布在跨膜区的突变位点中有46.2% (12/26)位于亲水性残基,由此推测,跨膜区的突变发生在亲水性残基的概率较大。杨中纯等(2010)人通过实验对GJB2蛋白跨膜区的突变R32H、V95L、R143W、S199F分别进行研究,结果表明R32H、R143W、S199F突变GJB2蛋白的表达缺失,V95M突变蛋白虽然在细胞膜上形成了间隙连接蛋白孔道却无生化偶联通透性的功能。Choi等(2009)人也通过实验证明跨膜区T86R突变使蛋白无法形成间隙连接功能;Ambrosi等(2010)人对跨膜区4种错义突变进行研究,发现其中T135A突变导致膜通道关闭,M34A突变导致膜通道的活性严重降低。此外,Oshima等(2011)人对M34A突变蛋白的电子晶体结构进行了研究,发现 M34A对N端通道口的形成产生影响,使通道几乎呈关闭状态。由此推测,当错义突变发生在跨膜区且突变改变了氨基酸的性质时,该突变将会影响蛋白在细胞膜上的表达或膜通道的通透性。
|
|
2讨论
间隙连接是细胞间直接进行物质交流的唯一通道,无论在脊椎还是无脊椎动物都发挥着重要的作用。在脊椎动物间隙连接的通道由connexin (Cx)蛋白家族构成,在小鼠体内约有20种间隙连接蛋白基因,在人类已知有21种间隙连接蛋白基因。所有的Cx都是具有4次横穿细胞膜的肽,形成了细胞外环、内环和跨膜区的三部分结构。跨膜区和细胞外区高度保守,然而不同Cx的功能差异是由这些区域的氨基酸序列的不同造成的。间隙连接介导的细胞间信号的异常可以引起全身多系统的疾病,由于不同的Cx在各器官、组织的分布不同,因此引起与之相关的疾病也不同。Cx26在全身大部分组织器官均有表达,并以皮肤和内耳表达水平最高。在内耳中,Cx26蛋白在人耳蜗的血管纹、基底膜、螺旋器均呈现较高水平的表达,GJB2基因的突变可致Cx26蛋白结构或功能发生障碍,从而导致遗传性NSHL。然而不同耳聋人群具有不同GJB2基因的突变谱(种族特异性),以及GJB2基因突变所致临床表型多样性,一直是该基因突变致聋研究的热点之一。
本文利用生物信息学方法对GJB2分子进化特征与NSHL的遗传效应进行了关联性分析,结果表明,人类GJB2氨基酸序列具有高度保守性,分析推测约有166个固定的氨基酸位点。GJB2序列的膜外区保守度高于跨膜区,跨膜区保守度高于胞质区,其中CT结构域保守度极低。序列109~125区域空间结构保守性低,两个非保守区位点发生致病性错义突变的概率较低。序列第113位和第123位推测为“空间结构保守位点”,预测发生在这两位上的错义突变会影响蛋白质的功能。发生在跨膜区错义突变如果改变了氨基酸的性质,则该突变将会影响蛋白在细胞膜上的表达或改变膜通道的通透性。参照Dai等(2009)对2 063例NSHL患者中发生了GJB2基因致病性错义突变的研究,从58例发生致病性错义突变的患者中得到10种错义突变,其中有1例患者同时发生了W3X和R143W突变,另1例患者同时发生Y152X和R127H突变。经比对,所检出的10种错义突变全部位于保守区,且发生在跨膜区的突变(R32C, S85P, T86R, R143W)均改变了氨基酸的性质,而在1号和2号非保守区均未发现致病性错义突变,这一研究结果也验证了保守性位点氨基酸的改变比非保守性位点更易致病,在非保守区出现致病性突变的可能性就较小。
通过对GJB2系统发育、三维结构及SOSUI分析,对GJB2分子进化特征有了进一步的理解,如我们首次推测并提出了人类GJB2序列第113位和第123位为“空间结构保守位点”的概念,并将继续予以验证。在对多物种序列比对中,我们发现绵羊与人类的序列比对得分较高(score=96),但两者空间结构存在一定的差异,这一结果与进化树中绵羊与人类的亲缘关系较远相一致。同样,褐家鼠与人类序列比对得分不如绵羊的高(score=93),但其三维结构却与人类几乎完全重叠,符合进化树中的亲缘关系。这些结果也进一步丰富了GJB2的生物信息学的资料。
目前,对GJB2致病突变性的研究大多采用基因序列分析、电子晶体结构分析、细胞与分子生物学等方法(Choi et al., 2009; Ambrosi et al., 2010; 杨中纯等, 2010; Oshima et al., 2011),而结合系统生物信息学方法的研究结果未见报道。本文利用生物信息学,对GJB2基因进行序列同源搜索,建立了该基因的物种序列库,对各物种基因进行多序列比对、系统发育与蛋白质三维结构空间比对分析,鉴定出序列保守位点和区域。并对保守位点(区域)、功能性位点(区域)和致病突变分布规律进行关联性分析,此为GJB2新致病性突变的预测以及突变致病机制研究提供了有用的前瞻性信息,同时也为其它疾病的相关研究提供了新的思路。
3材料和方法
3.1数据资料
从美国国立生物技术信息中心(NCBI)的数据库中,通过http//blast.ncbi.nlm.nih.gov/Blast.cgi查询获得17个物种的GJB2蛋白完整的氨基酸序列。
3.2 GJB2蛋白序列的生物信息学分析
3.2.1 ClustalW多重序列比对分析
用ClustalW2软件(www.ebi.al.uk/Tools/mas/clustalw2)对纳入研究的17个物种的GJB2蛋白序列进行多重序列比对分析(Thompson et al., 1994)。ClustalW是目前使用最广泛的多序列比对软件,它基于累进方法,采用两两比对、构建向导树、依次加入序列比对三个步骤进行多序列比对。比对时,先将多个序列两两比对构建距离矩阵;然后在根据距离矩阵计算产生系统进化指导树,按关系密切程度对序列进行加权;然后从最紧密的两条序列开始,逐步加入临近的序列并不断重新构建比对,直到所有序列都被加入为止。
3.2.2 GJB2的进化分析
用MEGA5.0软件绘制出纳入研究的17个物种的GJB2进化树,并且得到进化树中各结点处的祖先序列以供进一步研究(Jones et al., 1992; Tamura et al., 2011)。Mega是目前应用最广泛的进化分析软件之一,在本研究中采用了Maximum Likelihood法。
3.2.3蛋白质跨膜区与结构域分析
用SMART (http://smart.embl--heidelberg.de)对GJB2蛋白进行结构域功能分析SMART是能识别并注释蛋白质结构域的在线软件,它能对蛋白质的结构域进行研究和相互比较(Schultz et al., 1998;Letunic et al., 2009)。
3.2.4蛋白质跨膜区与结构域分析
用SOSUI对GJB2蛋白的跨膜区进行分析,SOSUI (http://bp.nuap.nagoya-u.ac.jp/sosui)是用于对膜蛋白进行识别分析并预测其跨膜螺旋轮结构的软件,其识别与预测精确度分别高达99%和97% (Hirokawam et al., 1998)。
3.2.5蛋白质三维同源模型构建与三维结构比较分析
用SWISS-MODEL (http://swissmodel.expasy.org/)对纳入研究的17条GJB2蛋白序列分别进行了同源建模(Guex and Peitsch, 1997; Schwede et al., 2003; Arnold et al., 2006)。然后,用VMD1.9软件(Board of Trustees of the University of Illinois and others)对序列与其对应的三维结构进行分析。蛋白质三维同源模型构建法是蛋白质三维结构预测最常见的方法之一,其原理是在蛋白质结构数据库中寻找未知结构蛋白质的同源构型,再将同源蛋白质的结构优化构建出预测的蛋白质三维结果。
作者贡献
范燚和喻岸竹完成数据处理、分析以及论文写作和修改。郁芸协助数据处理和部分修改工作。曹新在遗传性耳聋的数据分析和遗传效应分析方面给予了技术支持。韩新焕构思本研究、指导实施过程、修改并定稿本论文。全体作者均阅读了本文并同意论文全部内容。
致谢
作者感谢中国药科大学郑珩副教授的热情帮助。
参考文献
Ambrosi C., Boassa D., Pranskevich J., Smock A., Oshima A., Xu J., Nicholson B.J., and Sosinsky G.E., 2010, Analysis of four connexin26 mutant gap junctions and hemichannels reveals variations in hexamer stability, Biophys J., 98(9): 1809-1819 http://dx.doi.org/10.1016/j.bpj.2010.01.019 PMid: 20441744 PMCid: 2862186
Arnold K., Bordoli L., Kopp J., and Schwede T., 2006, The SWISS-MODEL workspace: A web-based environment for protein structure homology modeling, Bioinformatics, 22(2): 195-201 http://dx.doi.org/10.1093/bioinformatics/bti770
PMid: 16301204
Choi S.Y., Park H.J., Lee K.Y., Dinh E.H., Chang Q., Ahmad S., Lee S.H., Bok J., Lin X., and Kim U.K., 2009, Different functional consequences of two missense mutations in the GJB2 gene associated with non-syndromic hearing loss, Hum. Mutat., 30: E716-E727 http://dx.doi.org/10.1002/humu.21036 PMid:19384972
Dai P., Yu F., Han B., Yuan Y., Li Q., Wang G., Liu X., He J., Huang D., Kang D., Zhang X., Yuan H., Schmitt E., Han D., and Wong LJ., 2007, The prevalence of the 235delC GJB2 mutation in a Chinese deaf population, Genet. Med., 9(5): 283-289
http://dx.doi.org/10.1097/GIM.0b013e31804d2371 PMid: 17505205
Dai P., Yu F., Han B., Liu X.Z., Wang G.J., Li Q., Yuan Y.Y., Liu X., Huang D.L., Kang D.Y., Zhang X., Yuan H.J., Yao K., Hao J.S., He J., He Y., Wang Y.Q., Ye Q., Yu Y.J., Lin H.Y., Liu L.J., Deng W., Zhu X.H., You Y.W., Cui J.H., Hou N.S., Xu X.H., Zhang J., Tang L., Song R.D., Lin Y.J., Sun S.Z., Zhang R.N., Wu H., Ma Y.B., Zhu S.X., Wu B.L., Han D.Y., Wong LJ., 2009, GJB2 mutation spectrum in 2,063 Chinese patients with non syndromic hearing impairment, J. Transl. Med., 7: 26
http://dx.doi.org/10.1186/1479-5876-7-26 PMid: 19366456 PMCid: 2679712
Guex N., and Peitsch M.C., 1997, SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling, Electrophoresis, 18(15): 2714-2723 http://dx.doi.org/10.1002/elps.1150181505 PMid: 9504803
Hirokawa T., Boon-Chieng S., and Mitaku S., 1998, SOSUL: Classification and secondary structure prediction system for membrane proteins, Bioinformatics, 14(4): 378-379 http://dx.doi.org/10.1093/bioinformatics/14.4.378 PMid: 9632836
Jones D.T., Taylor W.R., and Thornton J.M., 1992, The rapid generation of mutation data matrices from protein sequences, Comput. Appl. Biosci., 8(3): 275-282 PMid: 1633570
Kelley P.M., Harris D.J., Comer B.C., Askew J.W., Fowler T., Smith S.D., and Kimberling W.J., 1998, Novel mutations in the connexin 26 gene (GJB2) that cause autosomal recessive (DFNB1) hearing loss, Am. J. Hum. Genet., 62(4): 792-799
http://dx.doi.org/10.1086/301807 PMid: 9529365
Letunic I., Doerks T., and Bork P., 2009, SMART 6: Recent updates and new developments, Nucleic Acids Research, 37(1): D229-D232 http://dx.doi.org/10.1093/nar/gkn808 PMid: 18978020 PMCid: 2686533
Mahdieh N., Rabbani B., Wiley S., Akbari M.T., and Zeinali S., 2010, Genetic causes of non syndromic hearing loss in Iran in comparison with other populations, J. Hum. Genet., 55(10): 639-648 http://dx.doi.org/10.1038/jhg.2010.96
PMid: 20739942
Martínez A.D., Acuña R., Figueroa V., Maripillan J., and Nicholson B., 2009, Gap-junction channels dysfunction in deafness and hearing loss, Antioxid. Redox Signal., 11(2): 309-322 http://dx.doi.org/10.1089/ars.2008.2138
Oshima A., Tani K., Toloue M.M., Hiroaki Y., Smock A., Inukai S., Cone A., Nicholson B.J., Sosinsky G.E., and Fujiyoshi Y., 2011, Asymmetric configurations and N-terminal rearrangements in connexin26 gap junction channels, J. Mol. Biol., 405(3): 724-735 http://dx.doi.org/10.1016/j.jmb.2010.10.032 PMid: 21094651 PMCid: 3026138
Schultz J., Milpetz F., Bork P., and Ponting C.P., 1998, SMART, a simple modular architecture research tool: Identification of signaling domains, Proc. Natl. Acad. Sci., 95(11): 5857-5864 http://dx.doi.org/10.1073/pnas.95.11.5857
Schwede T., Kopp J., Guex N., and Peitsch M.C., 2003, SWISS-MODEL: An automated protein homology-modeling server, Nucleic Acids Research, 31(13): 3381-3385 http://dx.doi.org/10.1093/nar/gkg520 PMid: 12824332 PMCid: 168927
Tamura K., Peterson D., Peterson N., Stecher G., Nei M., and Kumar S., 2011, MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods, Mol. Biol. Evol., 28(10): 2731-2739 http://dx.doi.org/10.1093/molbev/msr121 PMid: 21546353
Thompson J.D., Higgins D.G., and Gibson T.J., 1994, CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice, Nucleic Acids Research, 22 (22): 4673-4680 http://dx.doi.org/10.1093/nar/22.22.4673 PMid: 7984417 PMCid: 308517
Wu C.C., Hung C.C., Lin S.Y., Hsieh W.S., Tsao P.N., Lee C.N., Su Y.N., and Hsu C.J., 2011, Newborn genetic screening for hearing impairment: A preliminary study at a tertiary center, PLoS One, 6(7): e22314 http://dx.doi.org/10.1371/journal.pone.0022314 PMid: 21811586 PMCid: 3139636
Yang Z.C., Xiao Z.A., Xie D.H., and Xia K., 2010, Aberrant expression pattern of a novel mutation in connexin 26 gene resulting in autosomal recessive deafness, Zhonghua Yixue Yichuanxue Zazhi (Chinese Journal of Medical Genetics), 27(3): 241-245 (杨中纯, 肖自安, 谢鼎华, 夏昆, 2010, 连接蛋白基因一个新致聋突变体p.Y155X及功能分析, 中华医学遗传学杂志, 27(3): 241-245)