水稻休眠芽再生无性繁殖体系的构建 
2. 云南省高校滇型杂交粳稻分子育种重点实验室, 昆明, 650201
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2013 年, 第 11 卷, 第 6 篇 doi: 10.5376/mpb.cn.2013.11.0006
收稿日期: 2013年04月08日 接受日期: 2013年04月11日 发表日期: 2013年05月07日
引用格式(中文):
吕永刚等, 2013, 水稻休眠芽再生无性繁殖体系的构建, 分子植物育种(online), 11(6): 1032-1041 (doi: 10.5376/mpb.cn.2013.11.0006)
引用格式(英文):
Lv et al., 2013, The Establishment of Asexual Propagation System Based on Rice Dormant Buds Regeneration, Fenzi Zhiwu Yuzhong (online) (Molecular Plant Breeding), 11(6): 1032-1041 (doi: 10.5376/mpb.cn.2013.11.0006)
水稻休眠芽具有发育成完整水稻植株的潜能,是水稻再生繁殖的基础。本研究在头季稻收获后,截取植株地上茎节部分(含倒1~倒3节点休眠芽)为实验材料,比较通过激素处理、无激素处理、直接移栽稻桩以及种子萌发所获植株的各性状及生理指标差异。结果表明:不论是否通过激素处理,带有节点休眠芽离体稻杆的再生效率都极显著优于模拟再生稻而直接移栽的稻桩,其成苗率及再生植株的叶绿素含量、叶面积、穗长、单穗谷粒数等都大幅度提高。离体稻杆通过激素处理所获植株的各性状略好于无激素处理的再生植株,但与种子萌发的正常植株差异不显著。该研究初步建立了一套水稻休眠芽再生无性繁殖技术体系,能基本解决特殊材料的繁育以及现有再生稻产量不稳定和生产效率普遍偏低等难题,为丰富水稻无性繁殖技术提供了新思路。
水稻(Oryza sativa L.)是世界上重要的粮食作物之一,其产量直接影响世界粮食安全。要保障粮食产量稳步增长,就需要通过各种途径不断发掘新的种质资源,开拓新的生产技术,寻求经济而高效的生产模式。因此实现这一目标,就需在水稻的生产链上取得新的突破。再生稻的大面积推广、单基因控制的水稻雌性不育突变材料的发现和深入研究、新杂交制种模式的探索和提出,并以此探寻的新杂交育种模式等一系列研究都为我们提供了良好的发展思路。利用水稻带有节点休眠芽的离体稻杆进行育苗培养具有材料来源广,取材简单、快捷,而且结合外源植物激素使用和散播覆土培育方法简单和操作方便等优点,因而是一种在田间进行无性繁殖较理想的实验材料。1965年,Amemiya等开始水稻组织培养,几十年来,研究者先后从水稻各部位诱导出愈伤组织和再生植株(Nishi et al., 1968),目前用于水稻组织培养的外植体主要是成熟胚(阎丽娜等, 2010; 黄赛麟等, 2008)、幼胚(Hiei and Komari, 2008)、花药(Roy and Mandal, 2005)、幼穗(Shu et al., 2000; 王亚琴等, 2004)四种。但这些外植体研究目前尚未能完全脱离实验室,真正应用于实际生产的水稻类型或者生产上主推水稻品种的研究尚不多见,利用水稻上带有节点休眠芽的离体稻杆进行大田规模化育苗培养的研究未见报道。杨开渠首先利用水稻休眠芽的再生特性,开启了国内再生稻的生产研究(杨开渠, 1958, 农业学报, 9(2): 286-329)。研究表明头季稻稻桩倒1至倒3节点上的休眠芽芽都有较强的活力,表现为存活率高,干物重大等(施能浦, 1995),具有很大的再生潜力。为了提高再生效率和产量育种科学家借助一些外源植物激素对内源激素进行调节。植物激素对于植物的生长发育起着至关重要的作用,它几乎参与了植物生长发育的所有生理过程的调节:从细胞的生长、分裂和分化,到休眠、果实发育、性别分化和衰老及抗逆性等。有研究表明,休眠芽的萌发生长与内源激素有密切关系:内源细胞分裂素促进休眠芽的萌发(李经勇和唐永群, 2002),赤霉素(GA)、脱落酸(ABA)的含量和相互间的平衡也影响着休眠芽的萌发生长(禹利君等, 2008)。许多相关研究(Gubler et al., 2005;Rodríguez-Gacio et al., 2009)均指出,植物体内ABA的含量在解除休眠过程中有很大变化,cABA和fABA自由形式间存在逆反关系。当处于休眠状态时,ABA主要以自由形式(fABA)存在,而芽萌发时则多以结合形式(cABA)存在;一旦解除休眠进入生长状态以后,fABA所占的比例更少,cABA/fABA 值显著增加,这可能与fABA含量的减少、某些基因开启及DNA复制加强有关。研究还表明,植物激素参与调控水稻株高、分蘖、光合生产力等生长发育过程,而这些生长发育与水稻的产量有密切的关系(蔡传杰和陈善娜, 2001)。在双子叶植物的研究中发现,生长素(IAA)和细胞分裂素(CTK)在植物侧芽的生长过程中起关键作用(Chatfield et al., 2000; Shimizu-Sato and Mori, 2001)。Leopold(1949)认为禾本科作物同样受IAA主导的顶端优势影响,IAA抑制分蘖的发生,去除或削弱IAA活性可以解除顶端优势对分蘖生长的抑制。与IAA相反,外源施用CTK可以刺激分蘖的生长。但是,IAA可以抑制CTK对分蘖生长的促进效应(Anger et al., 1973)。水稻子粒中生长素含量与其灌浆过程密切相关,子粒中生长素主要在水稻灌浆前期起作用,并在很大程度上决定着水稻的维管发育状况、叶片光合能力及子粒灌浆速率,与水稻产量形成关系密切。吲哚乙酸(IAA)、吲哚丁酸(IBA)、奈乙酸(NAA)等外源激素对水稻增产效果十分明显。综上所述,以水稻上带有节点休眠芽的离体稻杆为材料,不仅可以充分利用休眠芽的再生潜能,而且还可实现完全脱离实验室在大田进行大规模的再生育苗,再结合一些外源植物激素使用还可以大大提高生产效率。但至今此方面的研究尚少,且相关繁育技术体系应用于实际生产还有相当的距离。
水稻雄性不育(male sterility, MS)和雌性不育(female sterility, FS)的发现及利用,杂交水稻的生产和发展都是保障粮食安全的重要举措。目前,在水稻杂交育种中,主要的技术体系是三系法和两系法,虽然这些体系都已较为成熟,但仍存在一些缺陷,主要体现在以下几点:(1)父母本分行或分区种植,开花时须人工赶粉辅助授粉,收获时父母本人工分收,费时费工;(2)制种产量低,不能很好解决种子混杂问题;(3)不适宜规模化、机械化操作等。基于此,可见杂交育种过程也需要耗费巨大的物力、人力以及漫长的育种周期。随着雌性全不育隐性单基因控制的水稻雌性不育突变体材料(fst)(由于胚珠和合子体分化发育异常导致完全败育,但雄性器官花粉发育正常)发现和在育种中的应用(Lee et al., 2013),利用雄性不育(MS)和雌性不育(FS),构建“MS+FS”的制种模式,即制种时的花粉供体为雌性不育系(陈丽娟等, 2009),也有望在不久后继三系和两系杂交的又一育种模式进行大规模的推广和应用。但问题在于其花粉供体是雌性不育系,不能正常结实获得大量的父本群体。因此,这种“MS+FS”制种模式能否得到成功应用,取决于能否获得大量的雌性不育株。还有一些特殊稀贵材料(如单体和三体以及多倍体, 自交不亲和等材料),由于自身的特点不能通过有性生殖的方式进行繁殖或不易繁殖。无性繁殖技术以及基因工程等是目前如何获得这些特殊材料最常用的方法途径。植物无性繁殖的方法中以组织培养在水稻中的应用最为广泛。但水稻组织培养也具有较大的局限性:幼胚、幼穗、花粉其取材受季节和环境的限制,取材时间严格,过早或过晚都会导致愈伤诱导失败;组织培养过程中容易出现死亡、白化等情况,难以获得大量的组培苗,且组培苗相对较弱,移栽到大田成活率得不到保障,同时对于大规模的生产成本高;组织培养目前也主要局限于实验室的研究,难以实现完全的室外生产。另一种被广泛应用的途径是利用水稻茎秆上的节点休眠芽再生进行无性繁殖,最典型的是再生稻的研究和推广。但目前再生稻仅限于在田间直接利用稻桩简单再生,不仅没有充分利用休眠芽,而且产量和生产效益也普遍偏低。通过如何合理挖掘水稻休眠芽的再生潜能,提高休眠芽的再生率和存活率以及再生植株的产量等方面入手,建立一套高效实用的无性繁殖体系还有待进一步深入研究。
本研究以水稻带休眠芽的离体稻杆为研究材料,通过数轮筛选适应范围较广的适宜外源植物激素组合及配比,对不同处理方式下获得再生植株的苗期、营养生长和生殖生长时期以及成熟期的各农艺性状和生理指标进行比较,阐述激素诱导离体休眠芽再生繁殖的可行性与高效性。本研究旨在克服现有水稻再生产量不稳定、产量和生产效益普遍偏低,以及特殊材料繁殖困难等生产和繁育技术难题,初步建立了一套完整的水稻休眠芽再生无性繁殖体系,不仅丰富了水稻无性繁殖的技术途径,同时也为“MS+FS”制种模式的应用推广奠定了基础。
1结果与分析
1.1适宜植物激素及组合配比筛选
1.1.1第一轮筛选结果
(1)激素单独使用时(表1),Kinetin浓度为9mg/L时出苗率最高为36%;NAA浓度为15 mg/L时出苗率达到38%,且随着浓度的升高出苗率还有增高的趋势;6-BA浓度为9 mg/L时出苗率最高为37% (图1A)。(2)组合激素使用时(表1),DK组合:0.1mg/L 2,4-D +(2.0~4.0) mg/L Kinetin,其出苗率均高于40% (图1B);NB组合:0.1 mg/L NAA +(7.0~12.0) mg/L 6-BA,出苗率均高于40% (图1B);KN组合:10.0 mg/L Kinetin +(1.0~6.0) mg/L NAA,出苗率均高于40% (图1A)。结果表明,植物激素组合使用的出苗率总体高于单独使用,这与王冬梅等(1996)、Rueb 等(1994)的研究结果一致,部分浓度区段效果较好,都可进一步筛选。
|
表1 第一轮筛选所用的激素、激素组合及浓度 Table 1 The first round of screening applied plant hormone, plant hormone combination and concentration |
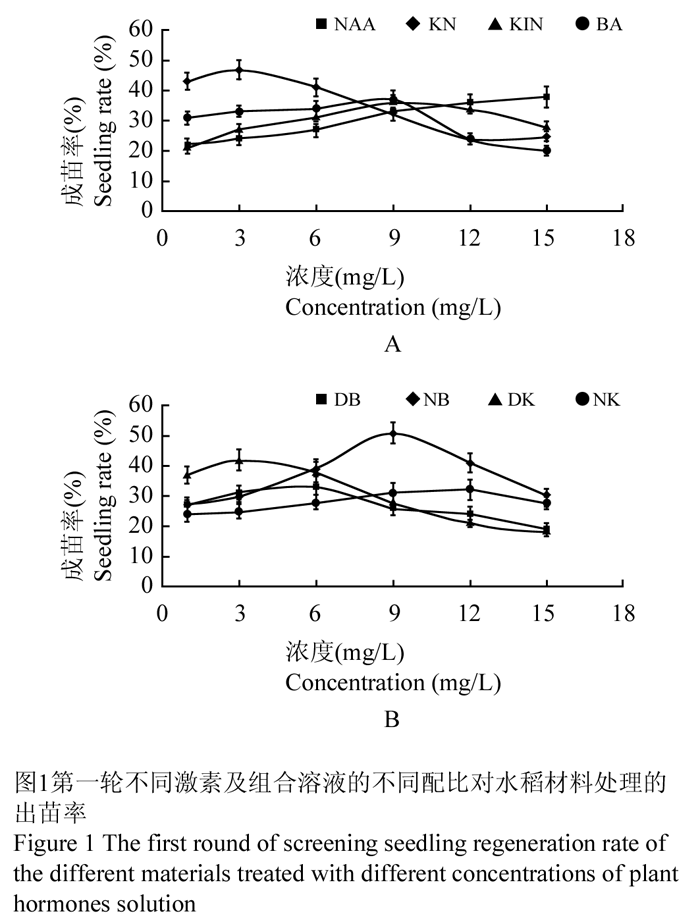 图1第一轮不同激素及组合溶液的不同配比对水稻材料处理的出苗率 Figure 1 The first round of screening seedling regeneration rate of the different materials treated with different concentrations of plant hormones solution |
1.1.2第二轮筛选结果
结果发现:DK(图2A)组合(0.1 mg/L 2,4-D + Kinetin),只有Kinetin浓度在(2.5~3.5) mg/L时D5A的出苗率高于40%,其余的出苗率都低于40%,结果均不理想;NB(图2B)组合(0.1 mg/L NAA + 6-BA),当6-BA浓度在(10.0~12.0) mg/L时,各材料的出苗率均高于40%,为适宜的植物激素组合配比;BN(图2C)组合(0.25 mg/L 6-BA + NAA),当NAA浓度在(30.0~60.0) mg/L时,各材料的出苗率均高于40%,为适宜的植物激素组合配比;KN(图2D)组合(10 mg/L Kinetin+NAA),当NAA浓度在(2.0~5.0) mg/L时,各材料的出苗率均高于40%,为适宜的植物激素组合配比。
综合FST、fst、D5A、黎榆B和滇杂37考虑:0.1 mg/L NAA+(10.0~12.0) mg/L 6-BA、0.25 mg/L 6-BA+(30~60) mg/L NAA、10mg/L Kinetin+(2.0~5.0) mg/L NAA出苗率较高,为适宜的植物激素组合配比,均能诱导带休眠芽的稻杆出苗,取得出苗率在40%~67%之间的好效果。
.png) 图2 第二轮筛选DK(A)、NB(B)、BN(C)、KN(D)溶液不同浓度处理的成苗率 Figure 2 The second round of screening seedling regeneration rate of the different materials treated with different concentrations of DK(A), NB(B), BN(C) and KN(D) solution |
1.2各材料不同处理的成苗率、干物重、穗长与穗粒数差异
总体上看(表2),各材料的离体稻杆通过植物激素处理,其成苗率极显著低于种子直接萌发(P<0.01),与无植物激素处理相比差异不显著(P>0.05),但极显著高于稻桩(P<0.01);不同处理的各材料分蘖盛期地上部分干物重,差异基本上不显著(P>0.05),只有用种子萌发所得D5A显著高于无激素处理(P<0.05)。此外,结果显示(表3)各材料通过植物激素处理的再生株穗长和穗粒数都极显著优于稻桩再生株(P<0.01),略好于无植物激素的再生株,但差异不显著(P>0.05),和种子萌发的植株相比差异也不显著(P>0.05)。表明带休眠芽的离体稻杆通过植物激素处理的成苗率明显不如用种子萌发的效率,但略好于无激素处理,极显著优于稻桩;不论是否通过植物激素处理还是种子萌发所获得的植株,其干物重差异都不明显;通过植物激素处理获得的植株穗长和单穗谷粒数都极显著优于稻桩直接再生的植株。
 表2 各材料不同处理的成苗率和干物重 Table 2 The seedling rate and dry weight of different varieties in different treatments |
 表3 各材料不同处理的穗长和穗粒数 Table 3 The panicle length and kernel number of different varieties in different treatments |
1.3各材料不同处理的叶绿素含量和叶面积差异
值得注意的是(表4),各材料的离体稻杆通过植物激素处理、无植物激素处理和种子萌发所获植株,其分蘖盛期所含的叶绿素含量和功能叶叶面积都基本上极显著优稻桩直接再生的植株(P<0.01)。而通过激素处理获植株的叶绿素含量和功能叶的叶面积都显著优于无激素处理所获植株(P<0.05),却种子发芽的植株没有明显差异(P>0.05)。表明离体稻杆通过植物激素处理所获植株的叶绿素含量及功能叶叶面积与种子萌发的植株差异不明显,但显著高于无激素处理所获植株和极显著高于稻桩再生植株。
 表4 各材料不同处理分蘖盛期的叶绿素含量、叶面积差异 Table 4 The leaf area and chlorophyll of different varieties in different treatments |
1.4初步建立完整的水稻休眠芽无性繁殖体系
经反复验证,本研究已初步建立了一套完整的水稻休眠芽无性繁殖技术体系(图3)。带有休眠芽的离体稻杆经筛选出适宜植物激素组合浸泡处理(图3A),之后散播于育苗棚或育苗床上覆土培育(图3B),待3叶~4叶期移栽至大田,同水稻常规大田栽培管理方法进行大田栽培管理就可获得大批量的再生植株(图3C~图3F)。表明以带休眠芽的离体稻杆为材料,经外源植物激素处理获得再生植株的繁殖技术体系有效、可行。
 图3 水稻休眠芽无性繁殖体系图解 Figure 3 Illustrated diagram of the asexual propagation system based on rice dormant buds |
2讨论
2.1构建水稻休眠芽再生繁殖体系的优势
水稻组织培养的特点限制了其难以挣脱实验室的束缚,实现大规模田间无性育苗。再生稻的发展虽然已有了相当的规模,但还没有充分挖掘和利用水稻休眠芽的再生潜能,且生产效率也普遍偏低。这些不仅是制约水稻再生育苗发展的瓶颈,也是主推和特殊水稻品种再生生产效率低下的主要限制因素之一。那么就急需找到一种适宜的室外培育材料和一套高效合理的再生繁殖技术体系。现已有不少的外植体被应用于组织培养(Mikami and Kinoshita, 1988),但适合于实验室外进行大规模再生繁育的培育材料,至今仍待继续探寻。植物的休眠芽作为一种优良的快繁材料在甘蔗的生产研究中应用的最为广泛也较成熟(曾吉恕, 1987, 作物杂志, 2: 32-33; 李富生等, 2000, 种子, (3): 41-45; Cheema and Hussain, 2004),它既可以用作原材料在实验室进行组织培养,也可以在田间直接掩埋上一季带有休眠芽的甘蔗杆来获得再生植株。在水稻研究中,利用休眠芽特性发展起来的再生稻研究也初具规模,但目前多局限于在田间直接利用稻桩进行简单的再生,再生效率和产量都面临严峻挑战。虽然也有不少的研究借助于植物激素的使用来增加产量和生产效率,但是也只是得到了小幅度的提升。因为其没有充分利用水稻不同节位上休眠芽、稻杆节点处的根发育系统、土壤养分的重复利用以及再生株生长空间狭小等。再生稻的生产利用虽然具有一定的局限性,但同时也具有相当大的发掘潜力。本研究利用带有休眠芽的离体稻杆培育,与稻桩直接再生相比,不仅大大提高了各节位休眠芽的存活率和再生率,而且穗部性状得到明显改善,每穗谷粒数也明显增多,因此说明若直接利用头季稻稻桩再生,其再生植株的生根区、叶形态建成和穗部发育可能都将受到营养供应等条件的限制,这与易镇邪等(2008)研究结果一致。
此外,本技术体系实施过程中仅需要少量的外源植物激素,相对于组织培养及三系和两系杂交制种大大降低了生产成本。以田间行、株距为15 cm×15 cm,每平方米单株种植49株为基准,本技术体系所用1亩头季稻稻杆的出苗量可供20~25亩移栽用苗所需,虽然相对于直接利用种子萌发移栽,其繁殖系数仍较低,但对于提高再生稻生产效率和一些用量较小的水稻材料繁育以及特殊材料的繁殖应用,特别是在“MS+FS”制种模式的应用推广中,此繁殖系数完全可以满足生产需求。因此,本技术体系若应用到水稻实际生产中,仍具有其可行性和高效性。
2.2提高水稻休眠芽再生繁殖体系生产效率的关键
本研究结果表明,将水稻休眠芽再生繁殖体系应用于生产,主要应考虑以下几个关键问题。其一,生育期的调整。这可以通过选择生育期较短的品种(组合)或提早播种期等方法来解决。其二,材料的整齐度问题。因为离体的稻杆包含不同节位的休眠芽,可能会出现同一批取材的休眠芽出芽先后不同,以致后期再生植株参差不齐。头季稻稻桩上不同节位干物质积累、碳水化合物和内源激素等含量存在显著差异(张桂莲等, 2005; 周文新等, 2008),结果会出现低位芽萌发早、生长缓慢,高位芽萌发晚但生长快,因此可通过利用不同外源植物激素和施加不同源的肥料来调节内源激素和库源来平衡休眠芽的出芽速率。也可以采取不同节位分别培育的方法来调整整齐度。其三,再生苗的成苗效率和成活率以及再生植株增产问题。水稻休眠芽的萌发与生长发育受植物激素的调控,使用外源植物激素可能会对内源激素的含量产生相应的影响,从而通过内源植物激素的变化调节休眠芽的萌发及再生植株的生长发育。本研究结果表明使用6-BA、NAA、CTK等不仅提高了成苗率,还提高了叶绿素含量、叶面积、穗长、穗粒数等主要指标,其原因可能是6-BA、CTK可以持续不断地刺激根芽细胞,使细胞增殖保持在最快阶段,促进节点处根的发育和再生苗的形成(Li et al., 2006)。然而现阶段对施用外源植物激素对内源激素的影响及与休眠芽萌发和生长关系的研究还较少,具体的影响机理还有待继续深入研究。
为完善水稻休眠芽再生繁殖技术体系的理论和实践,我们将继续深入研究施用外源植物激素对内源激素的影响机理;研究探明连续多代再生是否存在遗传力衰退的情况;实施品种配套再生繁育,为“MS+FS”制种模式的应用推广做好基础准备。
2.3离体节点休眠芽再生无性繁殖体系的应用前景
本研究表明,将头季稻带休眠芽的离体稻杆进行培育再生与稻桩直接再生的效果差异显著:前者主要充分利用了各个节位的休眠芽以及各节点处的根再生系统,且重新移栽可以通过控制移栽的密度来调控植株间的生长空间,再结合外源植物激素的应用大大提高了生产效率,因此该体系展现出了良好的发展潜力。在再生繁殖技术体系推广中,就是要方法简单方便高效,农民易接受。而本研究的技术方案不仅方法简单易懂,且高效成本低,因此,在一些特殊材料的繁殖,主推品种的再生生产以及未来杂交水稻“MS+FS”的新型制种模式中都具有很好的应用前景,将成为再生繁殖利用的主要途径。
3材料与方法
3.1试验材料
经筛选,本研究以水稻雌性核不育突变型品系(fs, female sterility)及其野生型品系(FST, FEMAIE STERILITY)、细胞质雄性不育系(滇粳优5号A (D5A))、常规品种(黎榆B)和杂种F1(滇杂37)各1份为试验材料。研究于2011年至2012年在云南农业大学(25.0°N, 102.7°E, 1895 masl.)和云南新平县戛洒镇(24.0°N, 101.3°E, 572 masl.)不同稻作生态试验田进行。
3.2材料的处理与移植
头季稻收获后7 d以内,截取供试材料带有倒1~倒3节点休眠芽的离体稻杆;稻杆洗净后散播在育苗床上并覆土,按水稻常规苗床育秧方法进行育苗和管理,或稻杆洗净后用植物激素组合物浸泡45~50 h,再散播在育苗床上并覆土,按水稻常规苗床育秧方法进行育苗和管理;待再生苗3叶~4叶期移栽大田,按水稻常规大田栽培管理方法进行大田栽培管理。
3.3适宜植物激素及组合配比筛选
3.3.1第一轮筛选
头季稻收获后7 d以内,截取供试材料带有倒1~倒3节点休眠芽的离体稻杆,洗净并等量混合各材料的稻杆,分别用不同浓度的植物激素及组合溶液浸泡48 h处理;将处理后的稻杆均匀散播在育苗床上覆土,按水稻常规苗床育秧方法进行育苗和管理;待再生苗3叶~4叶期分别统计出苗率,初筛出较好的激素及组合溶液配比。
3.3.2第二轮筛选
将获得的效果较好的激素及组合,方法同第一轮筛选,经第二轮筛选出适宜的组合及配比。
3.4测定项目
3.4.1考察出苗率、干物重、穗长和穗粒数
3叶~4叶期统计各材料稻桩、带休眠芽的离体稻杆以及种子萌发的出苗率。分蘖盛期分别取长势相当的各材料整株地上部分,先100℃烘箱中杀青1 h,然后置于80℃烘干至恒重,称重。成熟期考察穗长和单穗谷粒数。
3.4.2测定叶片叶绿素
分蘖盛期分别取各材料的新鲜叶片,去除叶脉后用电子分析天平精确称量0.05 g,剪碎后浸泡于装有80%丙酮的20 mL 容量瓶中,做3 次重复。密封置于4℃冰箱中浸提叶绿素约48 h 后,用80%的丙酮定容混匀,取浸提液用紫外可见分光光度计测定波长663 nm、646 nm 和470 nm 浸提液吸光值,按Lichtenthaler (1987)的方法,测定和计算叶片单位鲜重的叶绿素chla、chlb 和β-Car 的含量。
3.4.3测定叶面积
分蘖盛期分别取各材料的剑叶或倒二叶,按陶洪斌和林杉(2006)的打孔称重法,用孔径5 mm的单孔打孔器分别从距叶梢和叶柄2 cm处开始顺次打孔,每片叶片共打孔20次,并注意避开中心叶脉和已经枯萎的部分,将打下的圆形叶片计数并装入纸袋烘干(75℃下烘48 h)、称重(W1, 单位: g);打孔后的叶片装入纸袋烘干、称重(W2, 单位: g)。计算公式为:叶面积(cm2)=(W1+W2)×打孔数×r2×10-2/W1,其中:r为打孔器的半径,这里为2.5 mm。
3.5数据处理
数据处理和方差分析采用Microsoft Excel 和SPSS17.0 软件完成,图表中数据均为各处理的平均值±平均值标准误。
作者贡献
吕永刚和李东宣是本研究的实验设计和实验研究的执行人;吕永刚完成数据分析,论文初稿的写作;甘树仙、冯德党、董陈文华、熊海波、朱骞、张小玲、魏振飞、曹迎杰、张利东、伍腾飞参与部分大田试验及实验结果分析;陈丽娟和李东宣是项目的构思者及负责人,指导实验设计,数据分析,论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究得到了NSFC-云南联合基金重点项目(U1136604)、国家重点基础研究973项目(2011CB1-00401)和云南省基金重点项目(2006C006Z)共同资助。
参考文献
Anger R.H.M., Prasad P.C., Laude H.M., 1973, Effects of kinetin on tiller bud elongation in wheat (Triticum aestivum L.), Ann. Bot., 37(3): 565-571
Cai C.J., and Chen S.N., 2001, Advanced in studying of plant hormone, Yunnan Daxue Xuebao (Journal of Yunnan University), 23: 99-101,112 (蔡传杰, 陈善娜, 2001, 植物激素的研究进展, 云南大学学报(自然科学版), 23: 99-101, 112)
Chatfield S.P., Stirnberg P., Forde B.G., and Leyser O., 2000, The hormonal regulation of axillary bud growth in Arabidopsis, Plant J., 24(2): 159-169
http://dx.doi.org/10.1046/j.1365-313x.2000.00862.x
PMid:11069691
Cheema K.L., and Hussain M., 2004, Micropropagation of sugarcane through apical bud and axillary bud, International Journal of Agriculture and Biology, 6(2): 257-259
Chen L.J., Lee D.S., Li C.Y., Tan X.L., Xu X.Z., and Zhu Y.Y., 2009, Method of female sterility gene FST for hybrid rice breeding, China Patent, 200910094988 (陈丽娟, 李东宣, 李成云, 谭学林, 徐学洙, 朱有勇, 2009, 雌性不育基因FST用于杂交水稻育种的方法, 中国专利, 200910094988)
Gubler F., Millar A.A., and Jacobsen J.V., 2005, Dormancy release, ABA and pre-harvest sprouting, Curr. Opin. Plant Biol., 8(2): 183-187
http://dx.doi.org/10.1016/j.pbi.2005.01.011
PMid:15752999
Hiei Y., and Komari T., 2008, Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed, Nat. Protoc., 3(5): 824-834
http://dx.doi.org/10.1038/nprot.2008.46
PMid:18451790
Huang S.L., Lee D.S., Gan S.X., Zhu J.R., Li J., Liang J., and Chen L.J., 2008, Innovation of high effective regeneration system for matured embryo in vitro culture of rice, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 6(4): 801-806 (黄赛麟, 李东宣, 甘树仙, 朱建荣, 李娟, 梁晶, 陈丽娟, 2008, 水稻成熟胚培养高效再生系统的创新, 分子植物育种, 6(4): 801-806)
Lee D.S., Chen L.J., Li C.Y., Liu Y.S., Tan X.L., Lu B.R., Li J., Gan S.X., Kang S.G., Suh H.S., and ZhuY.Y., 2013, The Bsister MADS gene FST determines ovule patterning and development of the zygotic embryo and endosperm, PLoS One, 8(3): e58748
http://dx.doi.org/10.1371/journal.pone.0058748
PMid:23527017 PMCid:3602522
Leopold A., 1949, The control of tillering in grasses by auxin, American Journal of Botany, 36(6): 437-440
http://dx.doi.org/10.2307/2438077
PMid:18151120
Li J.Y., and Tang Y.Q., 2002, Relation of change in content of plant endogenous cytokinin to germination and growth of ratooning buds in hybrid rice, Zajiao Shuidao (Hybrid rice), 17(3): 50-52 (李经勇, 唐永群, 2002, 杂交水稻再生芽萌发生长与植株内源细胞分裂素含量变化的关系, 杂交水稻, 17(3): 50-52)
Li X., Mo X.R., Shou H.X., and Wu P., 2006, Cytokinin-mediated cell cycling arrest of pericycle founder cells in lateral root initiation of Arabidopsis, Plant Cell Physiol.,47(8): 1112-1123
http://dx.doi.org/10.1093/pcp/pcj082
PMid:16854941
Lichtenthaler H.K., 1987, Chlorophylls and carotenoids: pigments of photosynthetic membranes, In: Packer L., and Douce R. (eds.), Methods in Enzymology, Academic Press, New York, pp.350-382
Mikami T., and Kinoshita T., 1988, Genotypic effects on the callus formation from different explants of rice (Oryza sativa L.), Plant Cell, 12(3): 311-314
Nishi T., Yamada Y., and Takahashi E., 1968, Organ redifferentiation and plant restoration in rice callus, Nature., 219(5153): 508-509
http://dx.doi.org/10.1038/219508a0
PMid:5668441
Rodríguez-Gacio M.C., Matilla-Vázquez M.A., and Matilla A.J., 2009, Seed dormancy and ABA signaling: the breakthrough goes on, Plant Signal. Behav., 4(11): 1035-1049
http://dx.doi.org/10.4161/psb.4.11.9902
Roy B., and Mandal A.B., 2005, Anther culture response in indica rice and variations in major agronomic characters among the androclones of a scented cultivar, Karnal local, African Journal of Biotechnology, 4(3): 235-240
Rueb S., Leneman M., Schilperoort R.A., and Hensgens L.A.M., 1994, Efficient plant regeneration through somatic embryogenesis from callus induced on mature rice embryos (Oryza sativa L.), Plant Cell, Tissue and Organan Culture, 36(2): 259-264
http://dx.doi.org/10.1007/BF00037729
Shi N.P., 1995, Research and preliminary report about early hybrid cice-ratoon features and technology, Zajiao Shuidao (Hybrid rice), (1): 18-23(施能浦, 1995, 杂交早稻-再生稻栽培特性与技术研究初报, 杂交水稻, (1): 18-23)
Shimizu-Sato S., and Mori H., 2001, Control of outgrowth and dormancy in axillary buds, Plant Physiol., 127(4): 1405-1413
http://dx.doi.org/10.1104/pp.010841
PMid:11743082 PMCid:1540171
Shu L.H., Yin X.H., Ren X., and He G.C., 2000, Study on young panicle culture in vitro from wild rice of different genomes, Wuhan University Journal of Natural Sciences, 5(1): 119-122
http://dx.doi.org/10.1007/BF02828326
Tao H.B., and Lin S., 2006, Comparison on disc method with copy method and length-width method for measuring leaf area of rice, Zhiwu Shenglixue Tongbao (Plant Physiology Communications), 42(3): 496-498 (陶洪斌, 林杉, 2006, 打孔称重法与复印称重法和长宽校正法测定水稻叶面积的方法比较, 植物生理学通报, 42(3): 496-498)
Wang D.M., Huang X.L., and Huang S.Z., 1996, The action mechanism of cytokinins in plant tissue culture, Zhiwu Shenglixue Tongbao (Plant Physiology Communications), 32(5): 373-377 (王冬梅, 黄学林, 黄上志, 1996, 细胞分裂素类物质在植物组织培养中的作用机制, 植物生理学通报, 32(5): 373-377)
Wang Y.Q., Duan Z.G., Huang J.K., and Liang C.Y., 2004, Efficient regeneration from in vitro culture of young panicles of rice (Oryza sativa L.), Zhiwuxue Tongbao (Chinese Bulletin of Botany), 21(1): 52-60 (王亚琴, 段中岗, 黄江康, 梁承邺, 2004, 水稻幼穗培养高效再生系统的建立, 植物学通报, 21(1): 52-60)
Yan L.N., Li X., and Wu D., 2010, The comparison in tissue culture ability from mature embryo in different rice cultivars, Zhongguo Nongye Kexue (Scientia Agricultura Sinica), 43(6): 1127-1135 (阎丽娜, 李霞, 吴丹, 2010, 不同类型水稻材料成熟胚组织培养力的比较, 中国农业科学, 43(6): 1127-1135)
Yi Z.X., Wang X.H., Chen P.P., Zhou W.X., and Tu N.M., 2008, Differences in ratooning characteristics of axillary buds of hybrid rice with different ratooning types between in vivo and in vitro, Zajiao Shuidao (Hybrid Rice), 23(5): 73-76 (易镇邪, 王学华, 陈平平, 周文新, 屠乃美, 2008, 不同再生类型杂交稻腋芽在母体与离体条件下的再生特性差异, 杂交水稻, 23(5): 73-76)
Yu L.J., Shi Y.F., Xiao Y.H., Liu F.Z., Liu Z.H., 2009, Dynamic changes of endogenous GA3 and ABA contents in tea cultivars with different phenological characters and their impact on the regulation axillary buds sprouting, Zuowu Xuebao (Acta Agronomica Sinica), 34(2): 277-283 (禹利君, 史云峰, 肖海云, 刘富知, 刘仲华, 2008, 不同物候型茶树内源GA3和ABA的变化及其对腋芽萌发调控的影响, 作物学报, 34(2): 277-283)
Zhang G.L., Tu N.M., and Zhang S.T., 2005, Ratooning properties of axillary buds in hybrid rice, Zhongguo Shuidao Kexue (Chinese Journal of Rice Science), 19(4): 323- 327 (张桂莲, 屠乃美, 张顺堂, 2005, 杂交水稻腋芽再生特性, 中国水稻科学, 19(4): 323- 327)
Zhou W.X., Yi Z.X., Wang X.H., and Tu N.M., 2008, Difference in endogenetic hormone content of axillary buds from different nodes of two-line hybrid rice between in vivo and in vitro, Zajiao Shuidao (Hybrid rice), 23(3): 67-71 (周文新, 易镇邪, 王学华, 屠乃美, 2008, 母体与离体条件下两系杂交水稻不同节位腋芽的内源激素含量差异, 杂交水稻, 23(3): 67-71)