稻瘟病菌对非寄主植物毛竹的接种试验 
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2011 年, 第 9 卷, 第 8 篇 doi: 10.5376/mpb.cn.2011.09.0008
收稿日期: 2010年07月27日 接受日期: 2010年10月29日 发表日期: 2011年01月27日
郭小勤等, 2011, 稻瘟病菌对非寄主植物毛竹的接种试验, 分子植物育种 Vol.9 No.8 (doi: 10.5376/mpb.cn.2011.09.0008)
非寄主植物对非病原物所表现出的抗病形式是植物对致病微生物最普遍和持久的抗病形式,是最主要的抗病类型。本文探讨稻瘟菌的接种浓度和叶龄对侵染毛竹后的效果,并分析了毛竹对稻瘟病菌表现的非寄主抗性。研究结果表明,以1×106 cfu/mL浓度的稻瘟病菌接种毛竹嫩叶及水稻嫩叶后,水稻嫩叶发病严重,而所接种的稻瘟病菌分生孢子虽然能够在毛竹嫩叶中萌发形成附着孢,但细胞内菌丝数量很少。破坏了腊质层的毛竹叶面接种稻瘟病菌并不会形成过敏性坏死反应,推测可能是毛竹叶片腊质层中的障碍物赋予毛竹对稻瘟病菌的非寄主抗性。本研究结果可为进一步挖掘更多的抗稻瘟病菌基因提供理论基础。
植物总是处于许许多多病原微生物的包围之中,然而,每种植物都表现出对绝大多数病原微生物的抗性。植物表现出的这种对绝大多数病原微生物的抗性称作非寄主 抗性(Staskawicz et al., 1995; Heath, 1996)。相应地,一种植物对于大多数不能引起病害的微生物而言,称为非寄主。相反,针对那些有限的致病微生物而言则为寄主。基因对基因抗性是由来源于 植物的抗性基因(R)和来源于病原物的相应无毒基因(avr)之间的相互识别产生的(Flor, 1971; Bednarek and Osbourn, 2009; 阙友雄等, 2009),这种抗性比较容易被病原物所克服。与寄主抗性相比,非寄主抗性更广谱稳定,因此非寄主抗性基因在作物抗性改良中的作用倍受关注。非寄主抗性的 机制包括预存性结构屏障、预存毒性化合物如皂苷(主要作用于细胞膜中甾醇)以及由于病原菌侵染的诱导使植物产生的诱导型主动抗病反应(Alfano and Collmer, 2004; Nurnberger and Lipka, 2005; Ellis, 2006)。对非寄主抗性的研究有助于破译植物复杂的抗性防卫机制。
根据植物与病原物互作导致非寄主抗性时是否表现过敏性坏死反应 (HR),可将非寄主抗性分为两种类型,第一类非寄主抗性不能产生肉眼可见的坏死症状,第二类非寄主抗性产生HR,因此可见坏死症状(Mysore and Ryu, 2004)。非寄主植物所表现出的非寄主抗性类型取决于植物种类、病原菌种类,因此,某种非寄主植物对一种病原菌表现第一类非寄主抗性,而对另外一种病原 菌则可能表现为第二类非寄主抗性。迄今为止,围绕模式植物拟南芥(Lu, 2001; Collins et al., 2003; Kang et al., 2003; Shimada et al., 2006)、烟草(Hutcheson et al., 2001)、西芹(Kamoun, 2001)等,已经有多个不同的植物—病原菌互作系统被建立起来用于非寄主抗性的研究,并且在植物表现非寄主抗性的结构特征、生理生化机理、分子遗传规 律、信号传导、基因克隆等方面取得进展(Lipka et al., 2007; Bao et al., 2008; Kwon et al., 2008; Zellerhoff et al., 2010)。
毛竹与水稻同属禾本科植物,具有突出的抗水稻病虫害能力、基本不感染水 稻病虫害,若能从毛竹中挖掘到耐性基因,并转到水稻中,可增加水稻的可利用基因资源,有望获得高抗稻瘟病菌水稻。本文尝试建立毛竹—稻瘟病菌非寄主抗性互 作系统,从组织水平初步分析毛竹对稻瘟病菌表现的非寄主抗性,为挖掘毛竹中的抗性基因资源,应用于水稻的耐性改良奠定基础。这是竹类植物非寄主抗性研究的 首次报道。
1结果与分析
1.1稻瘟菌的接种浓度影响其侵染效果
由于尚未见到用稻瘟病菌菌株接种毛竹的相关报道,因此,在实验之前,首先对稻瘟病菌菌株07-73 (粳型)的接种浓度进行了探索。接种结果表明,接种浓度对毛竹产生过敏性坏死(HR)的程度有明显的影响。以浓度为1×105 cfu/mL的稻瘟病菌分生孢子接种后,毛竹的叶面并没有产生HR反应(图1A)。而以1×106 cfu/mL浓度的稻瘟病菌分生孢子接种时,毛竹叶面产生了强烈的HR斑(图1B)。表明稻瘟病菌菌株07-73 (粳型)刺激毛竹产生非寄主抗性的最适浓度以1×106 cfu/mL为宜。因此,以下的接种实验均选用了浓度为1×106 cfu/mL的稻瘟病菌菌株07-73(粳型)。
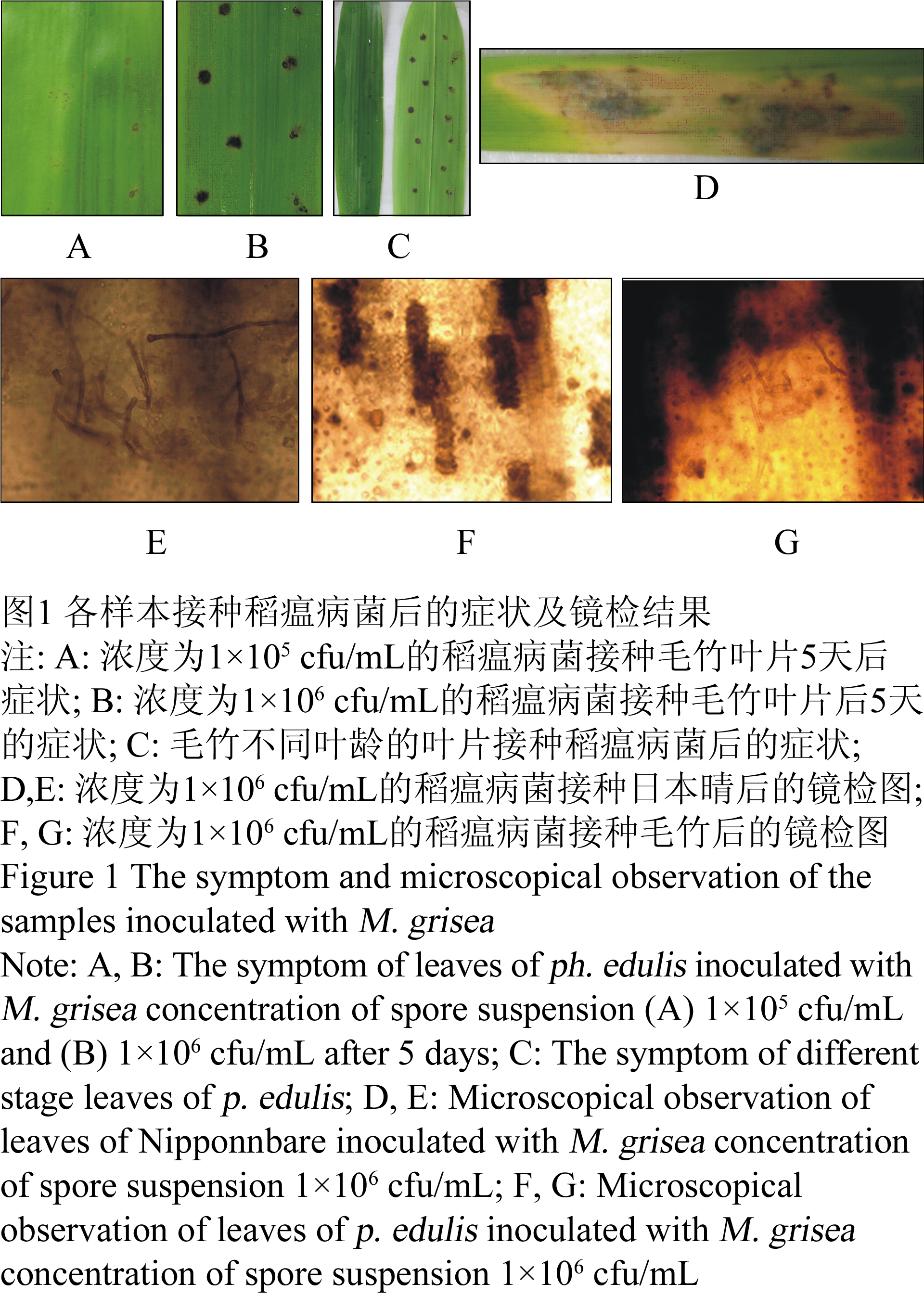 图1 各样本接种稻瘟病菌后的症状及镜检结果 Figure 1 The symptom and microscopical observation of the samples inoculated with M. grisea |
1.2非寄主植物毛竹的叶龄影响侵染效果
接 种实验表明,接种叶片的叶龄对非寄主抗性的产生也有很大影响。以同样浓度的孢子悬液对同不同叶龄的叶片进行接种后,发现,位于顶端的嫩叶对稻瘟病菌表现出 强烈的HR反应,而成熟叶却无反应(图1C中左是倒数第二片成熟叶,右为倒数第一片成熟叶)。因此,实验选用毛竹的嫩叶片进行接种。
1.3非寄主植物毛竹对稻瘟病菌表现抗性
以浓度为1×106 cfu/mL的孢子悬浮液分别接种日本晴和毛竹,结果发现,接种5天后,日本晴发病严重(灰色) (图1D)。40倍显微镜镜检观察到其体内产生了大量的稻瘟病菌菌丝和分生孢子(图1E)。然而以同样浓度的该菌接种的毛竹却对稻瘟病菌表现明显的非寄主抗性。在1×106 cfu/mL浓度下稻瘟病菌分生孢子刺激毛竹叶面产生强烈的过敏性坏死斑。在40倍显微镜下观察,发现在坏死部位有强烈的木质化现象(图1F),镜检时仅发现数例菌丝和孢子(图1G)。
1.4毛竹叶片的蜡质层阻碍了稻瘟病菌侵入
为 了验证是否是毛竹叶片表面的腊质层阻碍了稻瘟病菌的侵入和增殖,我们同时接种了破坏了腊质层的叶片和正常的叶片。试验结果表明,破坏了腊质层的叶片并不象 正常叶片那样能产生过敏性坏死斑(图1B右)。因此可以推测,很可能是毛竹叶片腊质层中的某种/些化学成分赋予了毛竹对稻瘟病菌菌株07-73(粳型)产 生的非寄主抗性。那么,究竟是什么物质使得毛竹对稻瘟病菌菌株07-73(粳型)产生了非寄主抗性呢,还需要进一步进行研究。
2讨论
稻 瘟病菌侵染寄主植物的一般过程为:孢子粘附在植物表面,然后萌发产生芽管,芽管伸长生长后形成附着胞;附着胞萌发生成侵染钉,穿透寄主组织继而在植物组织 内部定殖。在拟南芥与炭疽菌的非寄主抗性互作中,炭疽菌在拟南芥表面能形成黑变的附着胞,但不能在寄主细胞内进一步生长形成菌丝(Shimada et al., 2006)。与该结果相似,本研究发现,稻瘟病菌被接种到毛竹叶片后,能够萌发并形成黑变的附着胞。毛竹叶片接种部位会发生坏死,细胞内仅观察到很少的孢 子和菌丝的生长,表明,坏死部位限制了稻瘟病菌的发育,这一现象也存在于拟南芥与大豆锈菌的非寄主抗性互作中(Loehrer et al., 2008)。
有研究表明,来自寄主植物的物理和化学信号是激发稻瘟病菌分生孢子萌发和附着胞形成的重要因子。硬质、疏水的表面是促进稻瘟 病菌萌发的有利因素。抽提于寄主表面的蜡质能够促进附着胞的形成,而非寄主蜡质不能诱导分生孢子萌发和分化(Podila et al., 1993),但是分生孢子在感受这些化学物质之前需要有约2 h的对硬质叶面的感知过程(Hwang et al., 1995)。本研究发现被接种到毛竹叶片表面的稻瘟病菌能在24 h内发育形成附着胞,但是连接分生孢子和附着胞的芽管的长度很长。这些结果说明,稻瘟病菌在感受到毛竹硬质叶面后能够顺利地萌发生成芽管,但是由于作为非 寄主的毛竹叶面有不同于水稻的特殊化学物质和叶表面结构(地貌信息),对芽管的进一步分化发育不利,导致芽管持续生长,很难找到适宜的位置进行分化生成附 着胞,只形成少量的附着胞。这些结果也说明,毛竹叶面的某些“预存的”结构特征在其抵抗非寄主病原菌胶稻瘟病菌的过程中起了非常重要的作用,这种抗性即所 谓的“被动的防卫反应”(Mysore and Ryu, 2004)。本研究为采用遗传学和分子生物学等手段探索毛竹与稻瘟病菌之间的非寄主抗性中起关键作用的化学物质提供了基础。
3材料与方法
3.1植物栽培与真菌培养
水稻(Oryza sativa L.)品种为日本晴,由浙江省农业科学院植物保护与微生物研究所提供。种子经浸种(24 h, 28℃),催芽(48 h, 28℃)后播种在装有基质的直径10 cm塑料钵内。每钵播15粒发芽一致的种子,在相对湿度80%,16 h光照(27℃)、8 h黑暗(22℃)交替的培养室生长,稻苗长至5叶期时用于接种试验。试验前5 d酌施尿素一次。
毛竹(Phyllostachys edulis)种子来源于广西桂林,经浸种(24 h, 25~30℃),催芽(25℃, 95%)至胚根与种子等长时播种在穴盘中,在温室中待幼苗长到3~4片叶时,将其转移到小盆中,等苗长到5叶期时用于接种试验。
稻 瘟病菌:供试稻瘟病菌菌株07-73 (粳型),对日本晴表现强致病性,由浙江省农业科学院植物保护与微生物研究所提供,稻瘟病菌经燕麦片培养基(燕麦片50 g/L, 蔗糖20 g/L, 琼脂粉15 g/L)活化培养(28℃, 培养2周)后,接种于灭菌的大麦粒上,26℃培养10~14 d至菌丝体扩展至周围麦粒。产孢时,充分振动使麦粒均匀散开,用无菌水冲洗后将麦粒平铺于灭菌纱布上。室温条件下培养24 h,用无菌水洗下孢子,制成浓度为1×106孢子/mL的悬浮液,并添加Tween-20至终浓度0.05 %以供接种。
3.2接种
在 直径为9 cm的培养皿中铺上2层吸水纸,用水浸透后保湿,再将毛竹或水稻的叶片剪成长约4 cm的小段,放在吸水纸上,然后用蘸过水的手指轻轻擦一下毛竹或水稻叶片表面,使其均匀润湿,便于使孢子液滴留在叶片表面,取10 µL配制好的分生孢子悬浮液分别滴在水稻和毛竹叶片上,对照实验接种只含0.05% Tween-20的相同剂量无菌水溶液。接种后均置于暗室(25℃)保湿24 h,保持相对湿度近100%,然后重新放置培养室中以前述条件生长。5天后观察症状。在进行破坏蜡质层实验时,用牙签将蜡质层刺破,后续接种与上同。
3.3组织染色镜检
为 观察真菌的发育情况,对样本进行锥虫蓝(trypan blue)染色。将毛竹和水稻叶片剪成1.5 cm左右小块,置于Farmer氏溶液(乙酸: 乙醇: 氯仿=1: 6: 3, v/v/v)中透明化,然后在含0.05%锥虫蓝的乳酚与乙醇(1: 2, v/v)混合液中染色,再用水漂洗并浸于饱和水合氯醛中脱色。叶片放置于滴有50%甘油的载玻片上,用Zeiss Axioskop2 plus显微系统镜检,并用Zeiss Axiolam HRC相机组件拍摄图片。
参考文献
Alfano J.R., and Collmer A., 2004, Type III secretion system effector proteins: double agents in bacterial disease and plant defense, Annu Rev Phytopathol, 42(1): 385-414 doi:10.1146/annurev.phyto.42.040103.110731
Bao Y.M., Wang J.F., and Huang J., and Zhang H.S., 2008, Molecular cloning and characterization of a novel SNAP25-type protein gene OsSNAP32 in rice (Oryza sativa L.), Mol. Biol. Rep., 35(2): 145-152 doi:10.1007/s11033-007-9064-8
Bednarek P., and Osbourn A., 2009, Plant-microbe interactions: chemical diversity in plant defense, Science, 324: 746-748 doi:10.1126/science.1171661
Collins, N.S, Thordal-Christensen H, Lipka V, et al., 2003, SNARE-protein-mediated disease resistance at the plant cell wall, Nature, 425: 973-977 doi:10.1038/nature02076
Ellis J., 2006, Insights into nonhost disease resistance: can they assist disease control in agriculture? Plant Cell, 18(3): 523-528 doi:10.1105/tpc.105.040584
Flor H.H., 1971, Current status of the gene-for-gene concept, Annu Rev Phytopathol, 9: 275-296 doi:10.1146/annurev.py.09.090171.001423
Heath M C. 1996, Plant resistance to fungi, Can J Plant Pathol, 18: 469-475 doi:10.1080/07060669609500606
Hutcheson S.W., Bretz J., Sussan T., et al., 2001, Enhancer-binding proteins HrpR and HrpS interact to regulate hrp-encoded type III protein secretion in Pseudomonas syringae strains. Journal of Bacteriology, 183: 5589-5598 doi:10.1128/JB.183.19.5589-5598.2001
Hwang C S, and Kolattukudy P E. Isolation and characterization of genes expressed uniquely during appressorium formation by Colletotrichum gloeosporioides conidia induced by the host surface wax. Mol Gen Genet, 1995, 247: 282-294 doi:10.1007/BF00293196
Jones J D, Dangl J L. The plant immune system. Nature, 2006, 444(7117): 323-329 doi:10.1038/nature05286
Kamoun S. Nonhost resistance to Phytophthora: novel prospects for a classical problem. Curr Opin Plant Biol, 2001, 4(4): 295-300 doi:10.1016/S1369-5266(00)00176-X
Kang L, Li J, Zhao T, et al. Interplay of the Arabidopsis nonhost resistance gene NHO1 with bacterial virulence. Proc Natl Acad Sci USA, 2003, 100: 3519-3524 doi:10.1073/pnas.0637377100
Kwon C, Neu C, Pajonk S, et al. Co-option of a default secretory pathway for plant immune responses. Nature, 2008, 451(7180): 835-840 doi:10.1038/nature06545
Lipka V, Kwon C, Panstruga R, et al. SNARE-ware: the role of SNARE-domain proteins in plant biology. Annu Rev Cell Dev Biol, 2007, 23: 147-174 doi:10.1146/annurev.cellbio.23.090506.123529
Loehrer M, Langenbach C and Goellner K. Characterization of nonhost resistance of Arabidopsis to the asian soybean rust. MPMI, 2008, 21:1421-1430 doi:10.1094/MPMI-21-11-1421
Lu M, Tang X, and Zhou, J M. Arabidopsis NHO1 is required for general resistance against Pseudomonas bacteria. Plant Cell, 2001, 13: 437-447 doi:10.2307/3871287 doi:10.1105/tpc.13.2.437
Mysore K S, and Ryu C M. Nonhost resistance: how much do we know? Trends Plant Sci, 2004, 9(2): 97-104 doi:10.1016/j.tplants.2003.12.005
Nurnberger T, and Lipka V. Non-host resistance in plants: new insights into an old phenomenon. Mol Plant Pathol, 2005, 6(3): 335-345 doi:10.1111/j.1364-3703.2005.00279.x
Podila G K, Rogers L M, and Kolattukudy P E. Chemical signals from avocado surface wax trigger germination and appressorium formation in Colletotrichum gloeosporioides. Plant Physiol, 1993, 103: 267-272
Que YX, Song XX, Xu LP, Chen RK. Research progress on the interaction mechanism between plant and fungi. Letters in biotechnology. 2009, 20(2): 282-285 (阙友雄, 宋弦弦, 许莉萍, 陈如凯. 植物与病原真菌互作机制研究进展. 生物技术通讯, 2009, 20(2): 282-285)
Shimada C, Lipka V, O'Connell, R, et al. Nonhost resistance in Arabidopsis-Colletotrichum interactions acts at the cell periphery and requires actin filament function. Mol Plant Microbe Interact, 2006, 19(3): 270-279 doi:10.1094/MPMI-19-0270
Staskawicz B J, Ausubel F M, Baker B J, et al. Molecular genetics of plant disease resistance. Science, 1995, 268: 661-667 doi:10.1126/science.7732374
Zellerhoff N, Himmelbach A and Dong W B, et al. Nonhost resistance of barley to different fungal pathogens is associated with largely distinct, quantitative transcriptional respponses. Plant Physiol, 2010, 152: 2053-2066 doi:10.1104/pp.109.151829