水稻类辅助蛋白家族的生物信息学分析 
2.福建农林大学生物农药重点实验室, 福州, 350002
福建农林大学生物农药重点实验室,福州,350002
福建农林大学生物农药重点实验室,福州,350002
福建农林大学生物农药重点实验室,福州,350002
福建农林大学生物农药重点实验室,福州,350002
 作者
作者  通讯作者
通讯作者
基因组学与应用生物学, 2011 年, 第 30 卷, 第 34 篇 doi: 10.5376/gab.cn.2011.30.0034
收稿日期: 2011年06月03日 接受日期: 2011年06月24日 发表日期: 2011年07月11日
引用格式(中文):
王爱荣等, 2011,水稻类辅助蛋白家族的生物信息学分析,基因组学与应用生物学(online), Vol.30 No.34 pp.1218-1223 (doi: 10.5376/gab.cn.2011.30.0034)
引用格式(英文):
Wang et al., 2011, Bioinformatic analysis of the auxilin-like protein family in rice, Jiyinzuxue Yu Yingyong Shengwuxue (Genomics and Applied Biology), Vol.30 No.34 pp.1218-1223 (doi: 10.5376/gab.cn.2011.30.0034)
辅助蛋白作为热激蛋白Hsc70的伴侣分子在网格蛋白介导的细胞内吞作用中起重要作用。在水稻中还未发现有关类辅助蛋白的相关研究。为了明确水稻类辅助蛋白的编码基因、种类及其功能,本文对其进行了一系列的生物信息学分析。结果发现水稻中共有16个编码类辅助蛋白的基因,而且都有表达证据存在。这16个基因在水稻不同组织和不同胁迫条件下的的表达情况存在较大差异,说明它们所具有的功能不同。此外,进化分析发现,水稻类辅助蛋白共分为3个亚类,它们的分化早于单子叶植物和双子叶植物的分化。研究结果为进一步研究该类蛋白的分子和生物学功能奠定了良好基础。
辅助蛋白(auxilin)是第三类J蛋白家族中的成员,它的DnaJ结构域位于蛋白的C末端,而大部分J蛋白家族成员的DnaJ结构域都在N端(Walsh et al., 2004)。目前发现的辅助蛋白有两种类型,分别称为辅助蛋白1 (auxilin 1)和辅助蛋白2 (auxilin 2)。辅助蛋白1是动物大脑特异蛋白。辅助蛋白2在许多组织中都有表达,被称为与细胞周期G有关的蛋白激酶(cyclin G-associated kinase, GAK) ( Kanaoka et al., 1997; Kimura et al., 1997),它与辅助蛋白1有50%的相同性和60%的同源性(Eisenberg and Greene, 2007),除了N末端多一个能够磷酸化转录因子激活型蛋白1(activator protein-1, AP1)和2的α亚基的激酶结构域外,它和辅助蛋白1具有相同的结构域(Umeda et al., 2000; Korolchuk and Banting, 2002)。
辅助蛋白作为热激蛋白Hsc70的伴侣分子和Hsc70一起在细胞内吞作用循环的多个阶段中起着重要的伴护作用,如初生凹陷形成、网格蛋白包被凹陷(clathrin-coated pit, CCP)的内陷、CCP的溢缩以及CCP出芽以后网格蛋白包被囊泡(clathrin-coated vesicles, CCV)的脱包被等过程都离不开二者的相互作用(Eisenberg and Greene, 2007)。
在植物中,虽然有多个J蛋白已被研究证明参与了植物的不同信号途径如抗逆胁迫 (Zhu et al, 2010; Wang et al., 2004; Ham et al., 2006; Yang et al., 2010)、抗病途径(Shimizu et al., 2009)以及花信号途径(Shen et al., 2011)等等,但它们大多是第一、第二类型的J蛋白。关于辅助蛋白的研究非常少,研究较全面的只有拟南芥中的JAC1。Suetsugu等(2005)发现该基因所编码的蛋白可以在弱光条件下驱动叶绿体移动以有效获取光能;Takano et al.(2010)发现JAC1对叶绿体光定位移动的作用主要依赖于J结构域的活性;而Ezaki等(2007)发现该蛋白还具有不同的功能,它的过表达能够抑制根部对铝的吸收。我们的研究也发现拟南芥的另一个类辅助蛋白参与了抗病信号途径(宁正元等, 2007)。
水稻是重要的农作物之一,也是遗传育种、抗逆抗病机制研究的模式生物。目前,关于水稻J蛋白功能研究的报道非常少,Lu等(2009)发现在水稻中水稻条纹病毒的移动蛋白Pc4可以与水稻的两个J蛋白互作。Yamamoto等(2005)发现水稻的OsDnaJ 与增殖细胞核抗原蛋白OsPCNA互作,它在增殖细胞中强烈表达,且可被UV和破坏DNA的试剂如H2O2所诱导。Nguyen等(2004)发现了一个J蛋白被干旱胁迫所诱导。但这些J蛋白都不是类辅助蛋白家族。对于水稻中到底有多少个类辅助蛋白以及对它们的系统研究,目前尚未有报道。本文对水稻中的类辅助蛋白家族成员进行了一系列的生物信息学分析,为以后的功能研究奠定基础。
1结果与分析
1.1水稻中类辅助蛋白的鉴定及表达证据分析
通过保守结构域的搜索,并剔除DnaJ结构域在N末端的序列,结果发现水稻中有16个基因编码类辅助蛋白,但只有两个蛋白注释为辅助蛋白,其它的都注释为热激蛋白DnaJ (heat shock protein DnaJ)、DnaJ 同源蛋白亚家族C成员(DnaJ homolog subfamily C member)或含DnaJ结构域蛋白(DnaJ domain containing protein)。在Rice Genome Annotation Project数据库中搜索这16个基因的表达证据发现,只有Os07g28800没有表达证据(表1),但在MPSS中搜索基因在不同组织或不同胁迫条件下的表达证据,发现该基因只在干旱处理5d的14d幼苗根部和接种稻瘟病菌48 h 的敏感植物中有表达(图1, 3)。同时我们也搜索了拟南芥中的类辅助蛋白,发现拟南芥中有13个基因编码类辅助蛋白(表1)。
 表1水稻类辅助蛋白及其表达证据分析 Table 1 Auxilin-like proteins in rice and their expression evidence |
1.2类辅助蛋白的表达模式分析
为了探明水稻类辅助蛋白在不同组织和不同胁迫条件下的表达情况,我们在MPSS数据库中提取了水稻类辅助蛋白在不同组织及不同胁迫条件下的相对表达量并对其进行整理分析。结果发现,水稻类辅助蛋白在不同组织中的表达水平有所不同。其中仅在少数几个组织中能检测到的基因有:Os01g44310只在90 d未成熟的花序和35d的愈伤组织中有检测到,Os01g74580在14d的幼根和幼叶及萌发3d的种子中有检测到,Os03g10180在14d的幼叶、成熟花粉和90d未成熟的花序中有检测到,Os03g51830在14 d的幼叶、90d未成熟的花序和35d的愈伤组织中有检测到,Os07g28800在14 d的幼根和成熟花粉中有检测到,Os04g59060只在60 d的成熟干中有检测到,而Os01g16680没有在任何组织中检测到。其他几个水稻类辅助蛋白基因在多数组织中都有表达,但没有一个基因能在全部组织中都检测到,这说明水稻的类辅助蛋白存在组织表达特异性。在所有水稻类辅助蛋白和所有组织中,Os12g36180在60 d的顶端分生组织中具有最高表达水平(表2)。
 表2水稻类辅助蛋白在不同组织中的表达分析 Table 2 Transcript abundance of rice ALP genes in different tissues in MPSS libraries |
在非生物胁迫如盐、干旱和冷等条件下,水稻类辅助蛋白的表达水平各不相同。在高盐胁迫下,大部分基因都能在叶片中检测到,但仅有少数几个基因在根中检测到:Os01g25320、Os03g51830、Os05g33010和Os12g36180,也只有这四个基因可以同时在根和叶中有检测到,其中Os12g36180在叶中被诱导的最强烈。在干旱胁迫下,Os01g25320、Os03g51830、Os05g01590和Os12g36180可以同时在根和叶中有检测到,而Os01g16680、Os01g74580、Os04g59060、Os05g33010、Os07g03270在根和叶中都检测不到,其它几个只在根或叶一种组织中有检测到,而被干旱诱导最强烈的是Os03g51830。在冷胁迫下,所有基因在根中都没有检测到;而在叶中,有五个基因没有被检测到,它们是:Os01g16680、Os01g44310、Os02g10180、Os05g01590、Os07g28800。Os05g50370和Os12g36180表达量最高,也是在所有胁迫条件下及所有基因中表达量最高的(图1)。
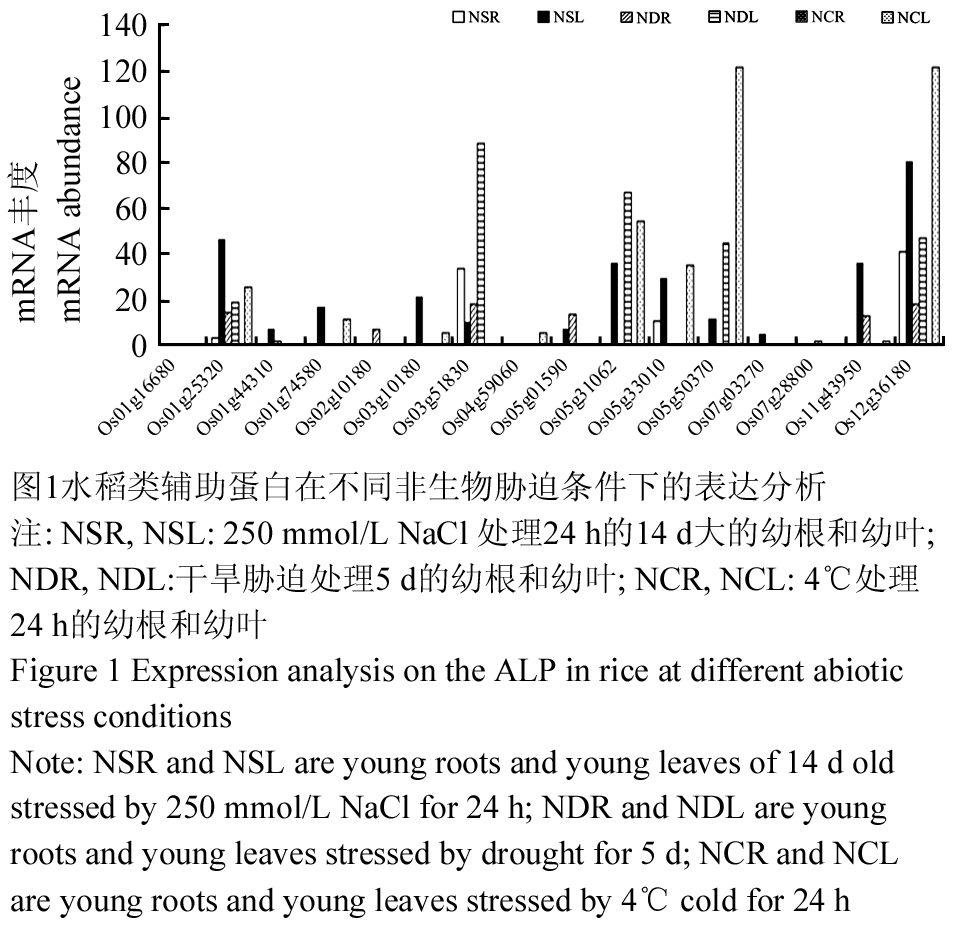 图1水稻类辅助蛋白在不同非生物胁迫条件下的表达分析 注: NSR, NSL: 250 mmol/L NaCl 处理24 h的14 d大的幼根和幼叶; NDR, NDL:干旱胁迫处理5 d的幼根和幼叶; NCR, NCL: 4℃处理24 h的幼根和幼叶 Figure 1 Expression analysis on the ALP in rice at different abiotic stress conditions Note: NSR and NSL are young roots and young leaves of 14 d old stressed by 250 mmol/L NaCl for 24 h; NDR and NDL are young roots and young leaves stressed by drought for 5 d; NCR and NCL are young roots and young leaves stressed by 4℃ cold for 24 h |
在生物胁迫如水稻接种Xanthomonas oryzae和Magnaporthe grisea后,水稻类辅助蛋白的表达水平也不同。在抗性品种和感性品种接种X. oryzae 12 h时Os3g10180被强烈诱导(图2),该基因在抗性品种中也被M. grisea强烈诱导(图3)。在接种X. oryzae和M. grisea的抗性植株和敏感植株的所有样品中,Os07g28800仅能在接种M. grisea 48 h的敏感植株中被检测到,而Os04g59060仅在接种X. oryzae 24 h后的抗性植株中被检测到(图2, 3)。在组织特异性和非生物胁迫相关的库中都没有检索到Os01g16680的表达信息,但在生物胁迫的表达库中有检测到。它在接种X. oryzae 6h和M. grisea 12 h的抗性植株中被强烈诱导表达(图2, 3)。
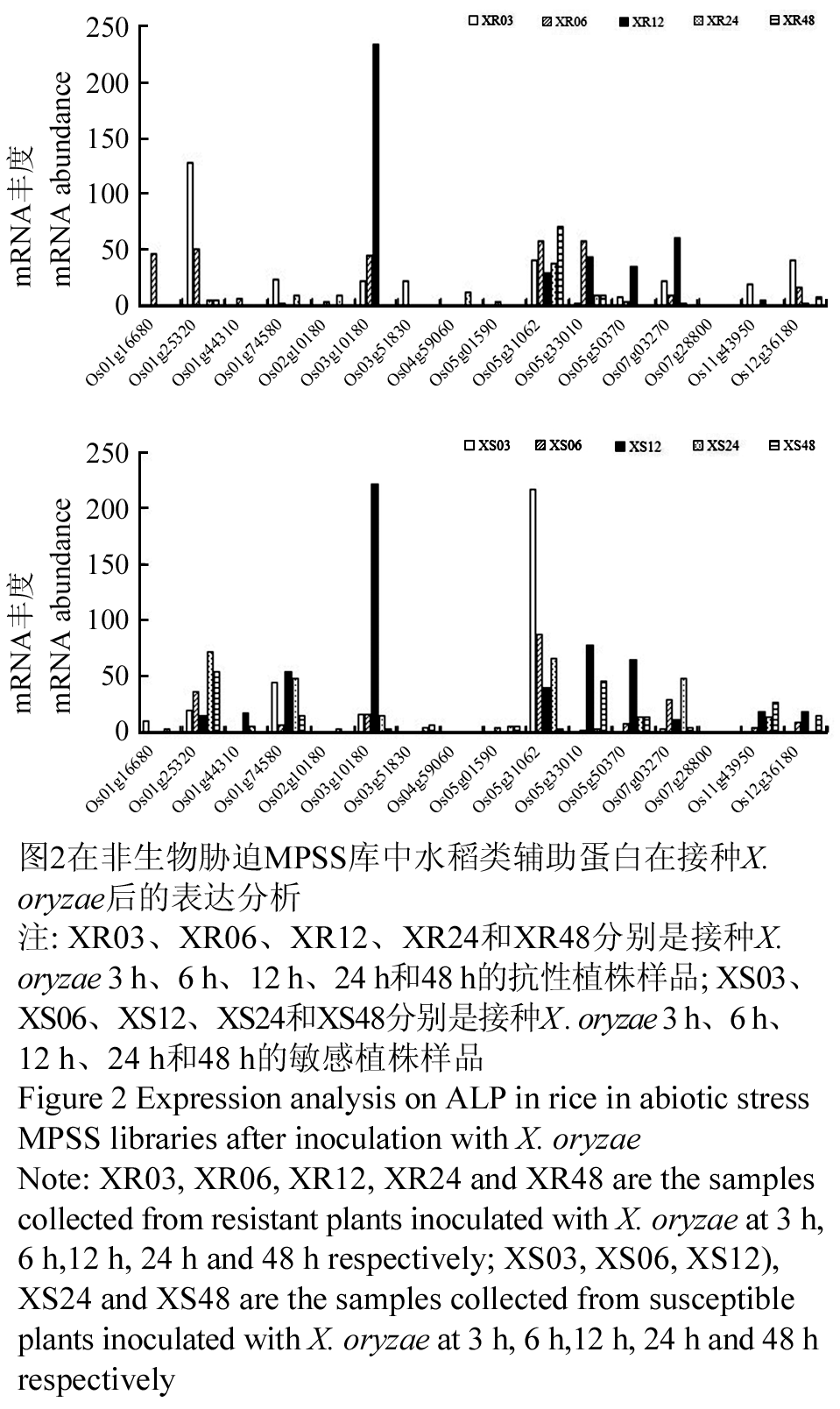 图2在非生物胁迫MPSS库中水稻类辅助蛋白在接种X.oryzae后的表达分析 注: XR03、XR06、XR12、XR24和XR48分别是接种X. oryzae 3 h、6 h、12 h、24 h和48 h的抗性植株样品; XS03、XS06、XS12、XS24和XS48分别是接种X .oryzae 3 h、6 h、12 h、24 h和48 h的敏感植株样品 Figure 2 Expression analysis on ALP in rice in abiotic stress MPSS libraries after inoculation with X. oryzae Note: XR03, XR06, XR12, XR24 and XR48 are the samples collected from resistant plants inoculated with X .oryzae at 3 h, 6 h,12 h, 24 h and 48 h respectively; XS03, XS06, XS12), XS24 and XS48 are the samples collected from susceptible plants inoculated with X. oryzae at 3 h, 6 h,12 h, 24 h and 48h respectively |
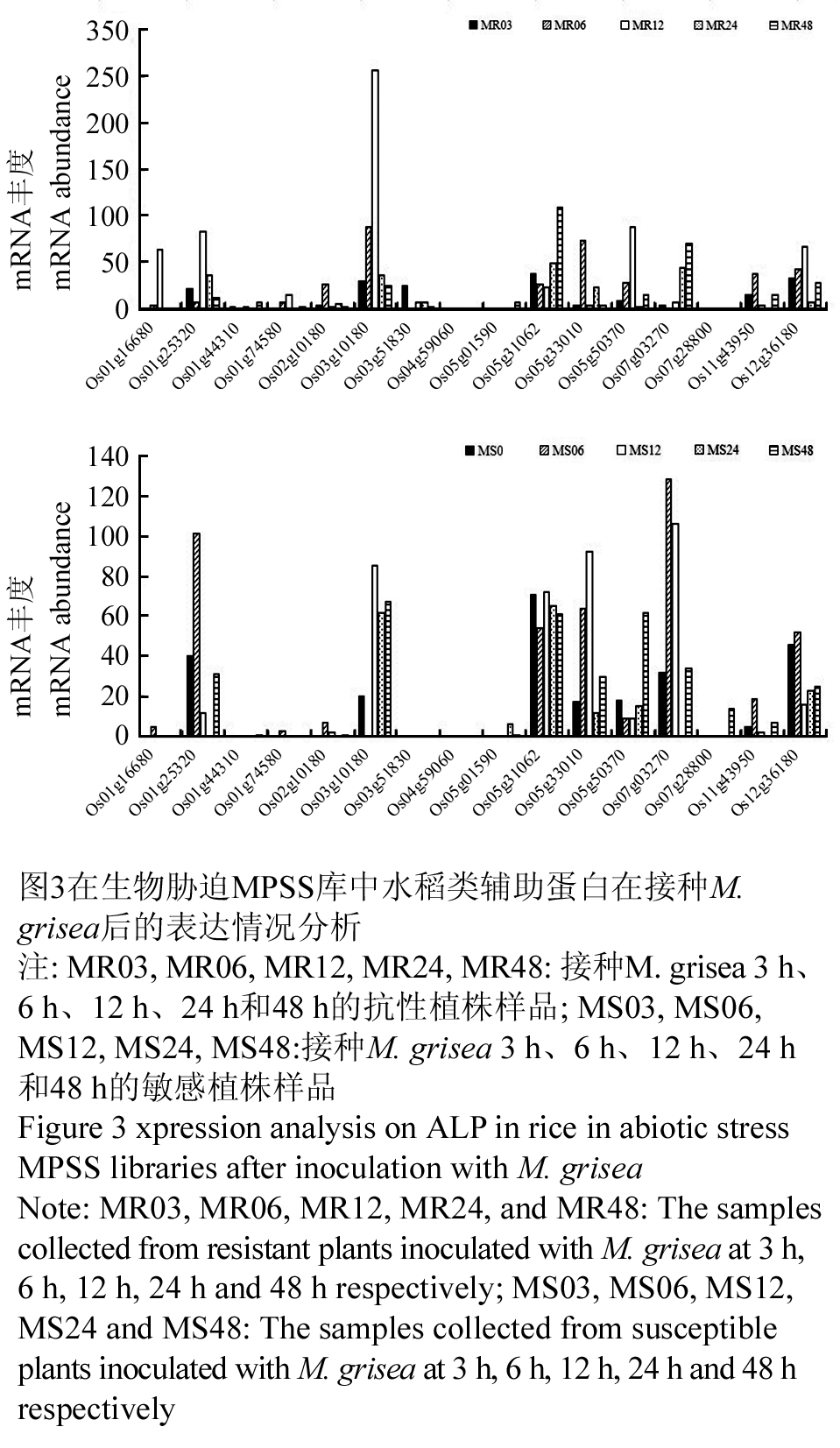 图3在生物胁迫MPSS库中水稻类辅助蛋白在接种M. grisea后的表达情况分析 注: MR03, MR06, MR12, MR24, MR48: 接种M. grisea 3 h、6 h、12 h、24 h和48 h的抗性植株样品; MS03, MS06, MS12, MS24, MS48:接种M. grisea 3 h、6 h、12 h、24 h和48 h的敏感植株样品 Figure 3 xpression analysis on ALP in rice in abiotic stress MPSS libraries after inoculation with M. grisea Note: MR03, MR06, MR12, MR24, and MR48: The samples collected from resistant plants inoculated with M. grisea at 3 h, 6 h, 12 h, 24 h and 48 h respectively; MS03, MS06, MS12, MS24 and MS48: The samples collected from susceptible plants inoculated with M. grisea at 3 h, 6 h, 12 h, 24 h and 48 h respectively |
1.3水稻类辅助蛋白的进化分析
为了分析水稻类辅助蛋白之间以及与拟南芥类辅助蛋白之间的亲缘关系,我们建了一个系统发育树。结果发现水稻类辅助蛋白和拟南芥的类辅助蛋白分为3个亚群:Os01g25320、Os01g44310、Os03g10180、Os05g50370、Os11g43950、Os12g36180和拟南芥的AT1G21660、AT1G30280、AT1G69060、AT1G75100、AT1G75310、AT4G12770、AT4G12780、AT4G36520、聚在一起,形成第一亚群;Os04g59060、Os05g01590、Os03g51830、Os05g33010、Os07g28800、Os07g03270和拟南芥的AT4G07990聚在一起形成第二亚群;Os01g16680、Os01g74580、Os02g10180、Os05g31062与拟南芥的AT1G09260、AT2G41520、AT5G03160、AT5G12430形成第三亚群(图4)。根据此系统进化分析可以推测,植物祖先基因组中的三类辅助蛋白在单子叶植物和双子叶植物分化之前就已经形成。
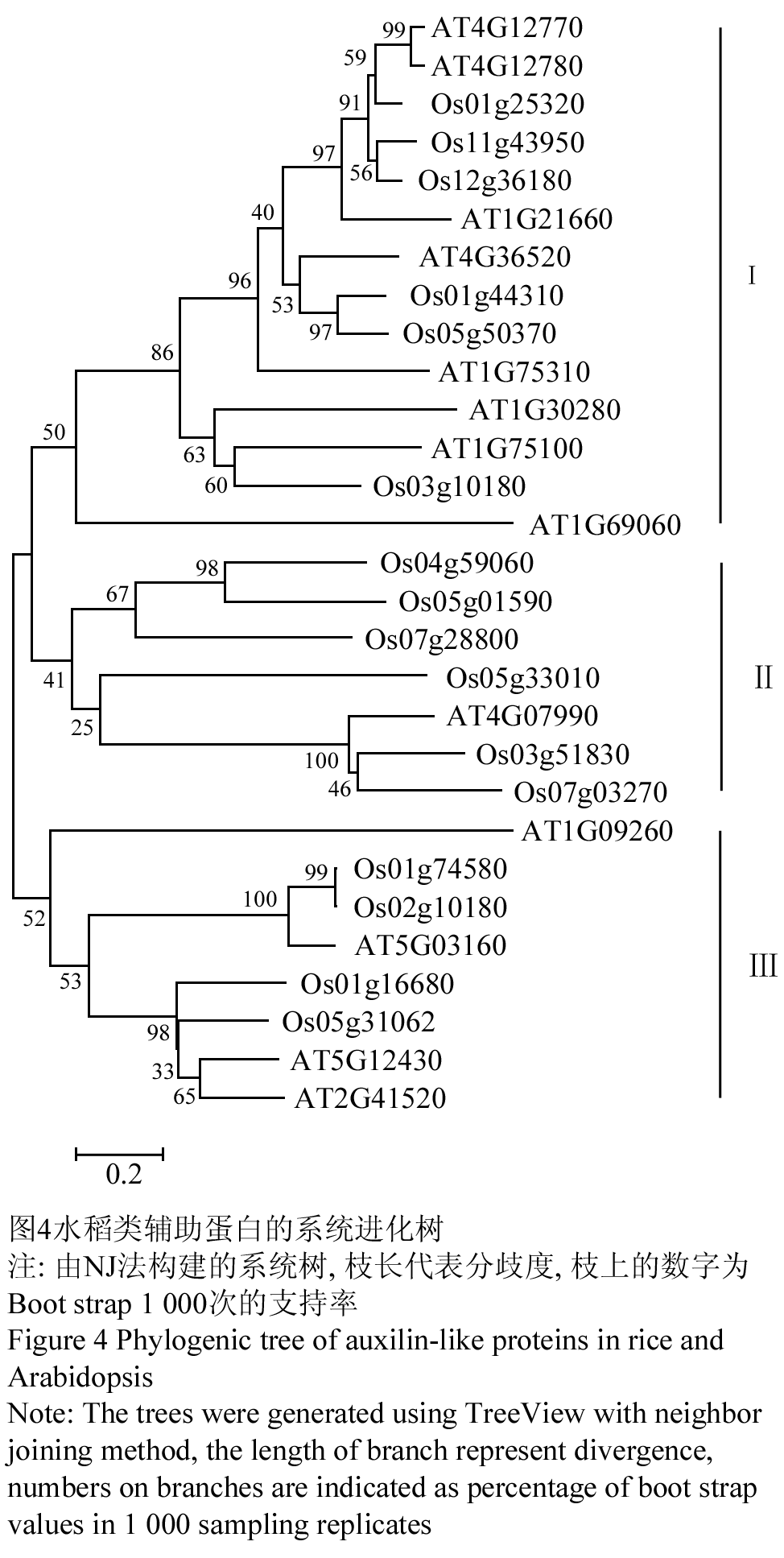 图4水稻类辅助蛋白的系统进化树 注: 由NJ法构建的系统树, 枝长代表分歧度, 枝上的数字为Boot strap 1 000次的支持率 Figure 4 Phylogenic tree of auxilin-like proteins in rice and Arabidopsis Note: The trees were generated using TreeView with neighbor joining method, the length of branch represent divergence, numbers on branches are indicated as percentage of boot strap values in 1 000 sampling replicates |
2讨论
本文首次运用生物信息学方法系统分析了水稻类辅助蛋白家族的结构、功能和表达特征。结果表明水稻全基因组中存在16个编码类辅助蛋白的基因,而且这些基因在不同组织、不同条件下的表达量不同,这说明不同类辅助蛋白所具有的功能不同。此外,同一组织中有多个类辅助蛋白表达,而且不同类辅助蛋白受同一胁迫的诱导,说明不同类辅助蛋白之间在调控上可能存在功能互补和功能冗余。Os12g36180在60 d的顶端分生组织、非生物胁迫如高盐和冷胁迫条件下是所有类辅助蛋白中表达量最高的,而在生物胁迫下表达量不高,说明该基因可能参与了植物的生长调控途径和抗非生物胁迫途径。与Os12g36180相比,Os05g50370在冷胁迫下的表达量也非常高,因此推测在冷胁迫下二者可能存在功能冗余或功能互补。Os03g51830被干旱诱导最强烈,说明它可能参与了抗旱信号途径。由于ABA是调控非生物胁迫包括盐、干旱和冷胁迫的重要信号分子(Koornneef et al., 1998; Leung and Giraudat, 1998; Finkelstein and Rock, 2002),因此Os12g36180、Os05g50370 和Os03g51830有可能参与了ABA信号途径。
在所有胁迫条件下,Os07g28800仅能在接种M.grisea 48h的敏感植株中被检测到,而Os04g59060仅在接种X.oryzae 24h后的抗性植株中被检测到,Os01g16680仅在接种X. oryzae 6h和M. grisea 12h的抗性植株中被强烈诱导表达(图2, 3),说明这三个类辅助蛋白分别参与了植物与病原菌的亲和互作与非亲和互作(Flor,1971; Valent and Chumley, 1991)。
DnaJ结构域是从酵母到哺乳动物的所有辅助蛋白中唯一一个保守的结构域(Xiao et al., 2006),而且是辅助蛋白与Hsc70相互作用的区域 (Jiang et al., 2003)。拟南芥的一个类辅助蛋白At4g12770已被证明可在体外使CCV脱包被(Lam et al., 2001)。水稻的类辅助蛋白也具有典型的DnaJ结构域,该类蛋白可能也与热激蛋白相结合,参与植物的胞吞胞吐作用。
3材料和方法
3.1数据库搜索
在水稻数据库(http://rice.plantbiology.msu.edu/analyses_search_domain.shtml)中输入DnaJ或其Pfam号:PF00226进行搜索,再用InterProScan (http://www.ebi.ac.uk/InterProScan/)和Pfam (http://pfam.sanger.ac.uk/)对所获得的蛋白氨基酸序列进行结构域分析,将DnaJ在N端的、没有DnaJ结构域及没有保守氨基酸HPD的蛋白除去。拟南芥类辅助蛋白的信息来源于TAIR数据库。
3.2蛋白表达分析
在http://rice.plantbiology.msu.edu/expression_expressed.shtml数据库中,输入编码类辅助蛋白的基因ID号,提交后得到基因表达的各种证据结果,然后对该结果进行进一步的统计分析。
在MPSS (http://mpss.udel.edu/rice/)数据库中,选择“17 bp signatures”后,输入基因的ID号获得水稻类辅助蛋白在不同组织及各种胁迫条件下的相对表达量。
3.3系统进化树分析
用Clustal x 1.83软件进行多序列比对,用Mega3.1软件、采用邻接法(Neighbor joining, NJ)构建系统发育树并进行1 000次Boot strap抽样。
作者贡献
王爱荣是本研究的实验设计和实验研究的执行人及论文初稿的写作;赵旭和林伟文参与实验设计与结果分析;鲁国东参与实验设计;王宗华指导实验设计,数据分析,论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由福建省科技厅青年人才项目(2007F3011)和福建农林大学青年教师科研基金项目(07A02)对本研究的共同资助。感谢两位匿名的同行评审人的评审意见和修改建议。
参考文献
Eisenberg E., and Greene L.E., 2007, Multiple roles of auxilin and hsc70 in clathrin-mediated endocytosis, Traffic, 8(6): 640-646
http://dx.doi.org/10.1111/j.1600-0854.2007.00568.x
PMid:17488288
Finkelstein R.R., and Rock C.D., 2002, Abscisic acid biosynthesis and signaling, In: Somerville C.R., and Meyerowitz E.M. (eds), The Arabidopsis Book, Published by American Society of Plant Biologists, Rockville, USA, pp.1-48
Flor H.H., 1971, Current status of the gene-for-gene concept, Annual Review of Phytopathology, 9: 275-296
http://dx.doi.org/10.1146/annurev.py.09.090171.001423
Ham B.K., Park J.M., Lee S.B., Kim M.J., Lee I.J., Kim K.J., Kwon C.S., and Paek K.H., 2006, Tobacco Tsip1, a DnaJ-type Zn finger protein, is recruited to and potentiates Tsi1-mediated transcriptional activation, The Plant Cell, 18(8): 2005-2020
http://dx.doi.org/10.1105/tpc.106.043158
PMid:16844903 PMCid:1533966
Jiang J., Taylor A.B., Prasad K., Ishikawa-Brush Y., Hart P.J., Lafer E.M. and Sousa R., 2003, Structure-function analysis of the auxilin J-domain reveals an extended Hsc70 interaction interface, Biochemistry, 42(19): 5748-5753
http://dx.doi.org/10.1021/bi034270g
PMid:12741832
Kanaoka Y., Kimura S.H., Okazaki I., Ikeda M. and Nojima H., 1997, GAK: A cyclin G associated kinase contains a tensin/auxilin-like domain, FEBS Letters, 402(1): 73-80
http://dx.doi.org/10.1016/S0014-5793(96)01484-6
Kimura S.H., Tsuruga H., Yabuta N., Endo Y. and Nojima H., 1997, Structure, expression, and chromosomal localization of human GAK, Genomics, 44(2): 179-187
http://dx.doi.org/10.1006/geno.1997.4873
PMid:9299234
Koornneef M., Léon-Kloosterziel K.M., Schwartz S.H., and Zeevaart J.A.D., 1998, The genetic and molecular dissection of abscisic acid biosynthesis and signal transduction in Arabidopsis, Plant Physiology and Biochemistry, 36(1-2): 83-89
http://dx.doi.org/10.1016/S0981-9428(98)80093-4
Korolchuk V.I., and Banting G., 2002, CK2 and GAK/auxilin2 are major protein kinases in clathrin-coated vesicles, Traffic, 3(6): 428-439
http://dx.doi.org/10.1034/j.1600-0854.2002.30606.x
PMid:12010461
Lam B.C.-H., Sage T.L., Bianchi F. and Blumwald E., 2001, Role of SH3 domain-containing proteins in clathrin-mediated vesicle trafficking in Arabidopsis, The Plant Cell, 13(11): 2499-2512
http://dx.doi.org/10.1105/tpc.13.11.2499
http://dx.doi.org/10.2307/3871590
PMid:11701884 PMCid:139467
Leung J., and Giraudat J., 1998, Abscisic acid signal transduction, Annual Review of Plant Physiology Plant Molecular Biology, 49: 199-222
http://dx.doi.org/10.1146/annurev.arplant.49.1.199
PMid:15012233
Ning Z.Y., Wang A.R., Lin W.W., Lin D.S., Zhang D.M., Liu H., Zhou J., and Wang Z.H., 2007, Bioinformatics and subcellular localization analysis of a protein containing the DNA_J domain of Arabidopsis thaliana, Fujian Nonglin Daxue Xuebao (Ziran Kexueban) (Jorunal of Fujian Agriculture and Forestry University (Natural Science Edition)), 36(4): 427-434 (宁正元, 王爱荣, 林文伟, 林德书, 张冬梅, 刘泓, 周洁, 王宗华, 2007, 拟南芥DNA_J结构域蛋白的生物信息学分析及其亚细胞定位验证, 福建农林大学学报(自然科学版), 36(4): 427-434)
Rajan V.B., and D’Silva P., 2009, Arabidopsis thaliana J-class heat shock proteins: Cellular stress sensors, Functional Integrative Genomics, 9(4): 433-446
http://dx.doi.org/10.1007/s10142-009-0132-0
Shen L., Kang Y.G., Liu L., and Yu H., 2011, The J-domain protein J3 mediates the integration of flowering signals in Arabidopsis, The Plant Cell, 23(2):499-514
http://dx.doi.org/10.1105/tpc.111.083048
PMid:21343416
Umeda A., Meyerholz A., and Ungewickell E., 2000, Identification of the universal cofactor (auxilin 2) in clathrin coat dissociation, European Journal of Cell Biology, 79(5): 336-342
http://dx.doi.org/10.1078/S0171-9335(04)70037-0
Walsh P., Bursać D., Law Y.C., Cyr D., and Lithgow T., 2004, The J-protein family: modulating protein assembly, disassembly and translocation, EMBO Reports, 5(6): 567-571
http://dx.doi.org/10.1038/sj.embor.7400172
PMid:15170475 PMCid:1299080
Xiao J., Kim L.S. and Graham T.R., 2006, Dissection of Swa2p/auxilin domain requirements for cochaperoning Hsp70 clathrin-uncoating activity in vivo, Molecular Biology of the Cell, 17(7): 3281-3290
http://dx.doi.org/10.1091/mbc.E06-02-0106
PMid:16687570 PMCid:1483056
Yang Y., Qin Y., Xie C., Zhao F., Zhao J., Liu D., Chen S., Fuglsang A.T., Palmgren M.G., Schumaker K.S., Deng X.W., and Guo Y., 2010, The Arabidopsis chaperone J3 regulates the plasma membrane H+-ATPase through interaction with the PKS5 kinase, The Plant Cell, 22(4): 1313-1332
http://dx.doi.org/10.1105/tpc.109.069609
PMid:20418496 PMCid:2879748