评述与展望
转基因技术的原理、方法与应用 
2诸暨市翠溪生物技术研究院, 诸暨, 311800
 作者
作者  通讯作者
通讯作者
欧洲生物技术研究进展, 2019 年, 第 9 卷, 第 1 篇
收稿日期: 2019年03月08日 接受日期: 2019年03月15日 发表日期: 2019年03月22日
Wu et al., 2019, Trangenesis: principle, technologies and application, Research Progress of Biotechnology in Europe, 9(1): 1-11 (doi: 10.5376/fbe.cn.2019.09.0001)
转基因技术作为现代生命科学的前沿技术之一,已经得到了稳定的发展和广泛的应用。转基因技术已经成为生物学家定向改造各种生物性状最为重要的手段。本综述详细阐述了转基因技术的原理,在动物、植物以及微生物等不同领域中的应用及其技术方法。随着转基因技术对人类生活影响的深入,迫切需要从学术和科普层面上,正确的引导及利用该技术,以使转基因技术更好的为人类发展而服务。
基因(gene)是遗传的物质基础,是DNA或RNA分子上含有特定遗传信息的一段核苷酸序列,是控制生物性状的基本遗传单位,是生命的密码,记录和传递着遗传信息。所有的基因都是由4种碱基组成,即A、G、T、C。我们可以知晓地球上的生物包括动物、植物、微生物数量巨大,种类繁多,形态各样这些都是有基因控制的,导致这些差异是由于基因的各个碱基的数量和编码形式不一样造成的。
基因能够通过复制把遗传信息传递给下一代,使后代出现与亲代相似的性状。人类的基因组中大约含有几万个不同的基因,这些基因储存着人类生命孕育、身体生长以及生命衰亡过程的所有信息,通过基因的复制、定向分化表达以及代谢异常基因的修复,完成生命繁衍、蛋白质合成以及组织和器官的分化生长等重要生理过程。可以说,生物体的生、长、老、病、死等一切生命内在和外在现象都与基因有关,基因也是决定人类身体健康的重要内在因素。
基因是转基因技术发展和应用的前提,而现代分子生物学是转基因技术的理论基础。转移的基因片段可以是从靶标生物体基因组提取具有特定性状的目的基因,也可以是通过人工合成的方法以获得特定序列的DNA片段。通过分子生物学的方法,将目的基因的DNA片段转入到特定的生物中,使其与特定生物的基因组进行重组,再从获得的重组体群中进行多代的人工选育过程,从而获得具有稳定的特定遗传性状的个体。转基因技术可以按照人类的意愿对动物、植物和微生物等进行定向的改造,进而培育出新品种。
1974年,科恩将金黄色葡萄球菌质粒上所携带的抗青霉素基因成功的转移到大肠杆菌体内,从而揭开了转基因技术商业化应用的序幕(Chang and Cohen, 1974)。发现DNA限制酶的纳森斯、亚伯与史密斯获得了1978年的诺贝尔医学奖,当时斯吉巴尔斯基在《基因》期刊中写道:限制酶将带领我们进入合成生物学的新时代(Hinnen et al., 1978)。
1982年,美国Lilly公司成功实现了利用大肠杆菌生产重组胰岛素的目的,这也标志着世界上第一个基因工程药物的诞生。1992年荷兰科学家成功培育出能够产人促红细胞生成素的转基因牛,而人促红细胞生成素在人体中具有能刺激红细胞生成的功能,是目前治疗贫血的良药。随着转基因技术的不断发展,该技术在药物生产中逐渐凸显出重要的利用价值。
20世纪,转基因技术的出现,标志着通过基因工程技术能够对不同种类生物的基因进行重组,而人类也可以根据自己的意愿定向地改造生物的遗传特性,进而创造出新的生命类型和物种(Ledford, 2016)。数十年来,通过转基因技术产生的新的农作物和动物品种数以千计,其在农作物品种改良、疾病防治、新型药物生产方面产生巨大推动作用,逐渐影响着人类的生产和生活。
1转基因的原理
转基因技术的原理是通过分子生物学技术,人为的分离或修饰具有特定遗传性状的优质目的基因,将其通过特定途径导入到靶标生物体的基因组中,由于导入基因的表达会引起靶标生物体的特定性状发生可遗传的修饰改变,从而达到改造生物特性、获得新的生物体的目的,这种技术称之为人工转基因技术(transgene technology)。
人工转基因技术常用的方法和工具包括基因枪、电破法、显微注射和脂质体等,转基因技术最初是用于研究某一特定基因的功能,通过把外源基因以某种途径导入到受体生物体(一般为模式生物, 如拟南芥或斑马鱼等)的基因组并发生有效重组,通过观察重组生物体所表现出的性状,进而达到揭示特定基因功能的目的。
转基因技术按照技术途径的不同,主要可以分为自然转基因和人工转基因2大类;按照研究对象可分为植物转基因技术、动物转基因技术和微生物基因重组技术这3类。
1.1人工转基因技术
目前学术界或者民间流传的的“基因工程”、“遗传工程”或“遗传转化”等术语都是转基因的同义词。从狭义层面上来说,能够改变动植物性状的人工技术都可以被称为转基因技术,而对微生物的遗传性状改变的操作则被称为遗传工程技术。目前媒体上被称为“遗传修饰过的生物体”(genetically modified organism, 简称GMO)通常是指经转基因技术修饰过的生物体。
基因工程是在分子水平上按照人们的意愿对生物进行改造,技术要求高,过程复杂,又是一个有机的整体,人工转基因主要步骤是“获取目的基因”、“重组DNA”、“目的基因的检测与表达”(Shou et al., 2002) (图1)。
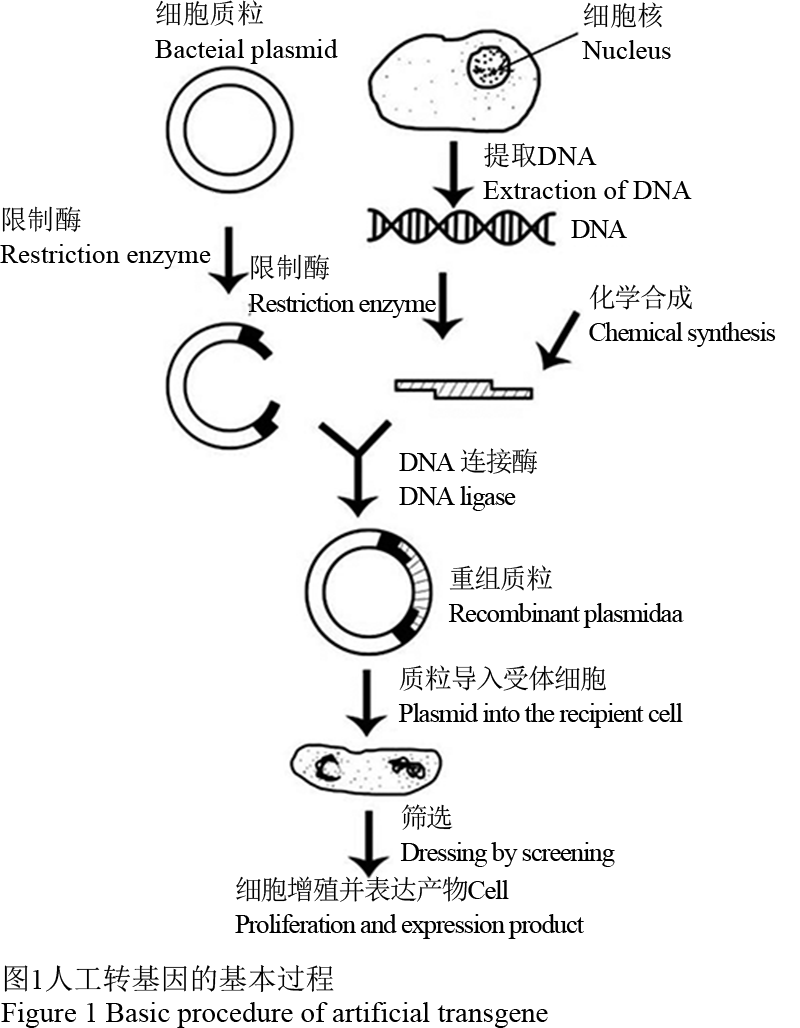 图 1 人工转基因的基本过程 Figure 1 Basic procedure of artificial transgene |
自然转基因是指自然界中动物、植物或微生物自主、天然发生的基因转移的现象,不是人为导向的行为,例如乙型肝炎病毒DNA通过自然感染的途径被整合到人精子细胞染色体上(周旭平, 2011)、噬菌体将自己DNA的插入到溶源细胞DNA上等等自然现象。
自然发生的转基因事件在自然界中普遍存在,而基因组是否能够取得新基因进而发生重组的重要途径是外源基因的获得,这也是生命进化、物种演化过程中的一个非常重要的推动力量(Arias and Rieseberg, 1994)。例如:科学家发现自然环境下(预计早在8000年前),土壤农杆菌通过Ti质粒侵染红薯,将T-DNA转移给了红薯,结果发现红薯发生了意想不到的有利突变,人类通过持续的选育获得更优良的红薯品种,并食用了长达数千年。
1.3植物转基因技术
植物转基因技术可以通过染色体工程技术、遗传物质转移、原生质体融合和细胞重组等技术途径实现将外源目的基因导入靶标植物,通过改变植物的某些遗传特性,进而培育出产量高、品质优、耐储藏等新特性的作物品种,同时还可以实现植物抗病毒、抗寒、抗旱、抗盐碱、抗虫和抗除草剂等特性,如抗虫棉、抗除草剂大豆、北极鳄梨(Berge, 2012)、转基因三倍体毛白杨等。而且可以将转基因植物或离体培养的细胞作为“工厂”,来生产导入的外源基因的表达产物,目前已商业化的主要有人的生长激素、干扰素、胰岛素、表皮生长因子、白介素2和乙型肝炎疫苗等产品,均已实现了植物体表达或者细胞工厂生产。
1983年世界第一例转基因植物——烟草问世以来仅30多年的时间(图2),转基因植物的研究和应用获得了巨大的发展和成果。
.png) 图 2 世界上第一例转基因植物——烟草 Figure 2 The world's first transgenic plant, tobacco |
1.4动物转基因技术
动物转基因技术就是将外源目的基因导入靶标动物的基因组中的技术。按照技术方案设想,其可以通过细胞重组、细胞融合、基因工程和染色体工程等技术将外源目的基因导入受体动物的卵细胞、精子或受精卵中,再通过生殖技术进行选育,进而培育成所设计的转基因动物。
世界上第一个被成功克隆的哺乳动物——克隆羊多莉(图3),就是由移植母羊的乳腺细胞到被摘除细胞核的卵子细胞中发育而成的。它证明了一个哺乳动物的特异性分化的细胞也可以发展成一个完整的生物体。
.png) 图 3 克隆羊多莉 Figure 3 Dolly the sheep |
动物生长素基因、高泌乳量基因、多产基因、瘦肉型基因、抗寄生虫基因以及抗病毒基因等特殊功能性基因是影响动物生长发育的关键基因,通过对这些功能性基因的利用,可以培育成生长周期短、产仔量多、泌乳量和蛋白含量高的转基因动物新品种。同时还可以促使转基因动物新品种其所生产的肉类口感、加工性能和皮毛品质更好,并使新品种动物具有抗病性,提高生存效率,目前转基因动物技术已经在牛、羊、猪、鸡、鱼等家养经济类动物中取得了显著的成果。
1988年,澳大利亚scammark教授通过基因工程技术将猪生长激素基因注入到猪的受精卵中,成功的获得了“超级猪”(图4),这是转基因动物技术在家畜基因工程育种领域里的一大突破。
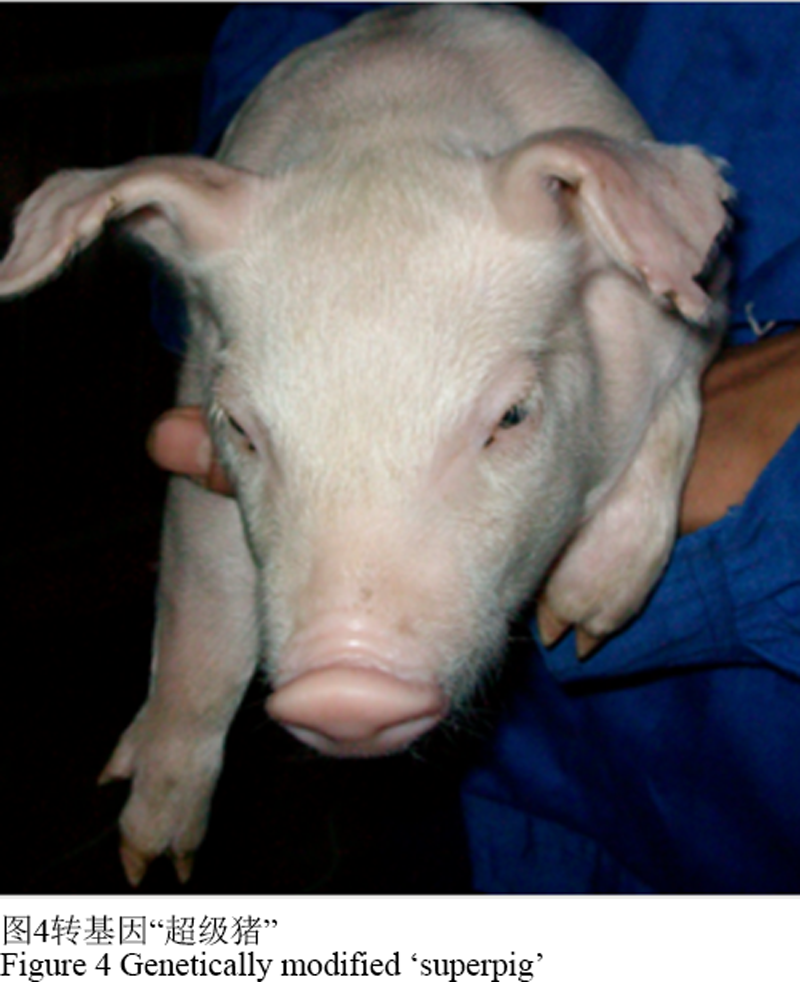 图 4 转基因“超级猪” Figure 4 Genetically modified ‘superpig’ |
但由于受到遗传镶嵌性和杂合性的影响,转基因动物其有性生殖后代通常存在很大的变异,难以形成能够稳定遗传的转基因动物品系。因而,科学家们尝试将外源目的基因导入线粒体进行重组,形成稳定的遗传体系,再将重组质粒送入靶标动物的受精卵中,进而筛选可稳定遗传的转基因动物品系。科研事实证明,由于线粒体的细胞质遗传特性,通过该途径育成的动物其有性后代基本都是转基因个体,而且能够进行稳定的遗传。
1.5微生物重组技术
微生物基因重组技术是目前所有转基因技术中应用最为广泛的技术领域,与动物和植物转基因技术不同的是,微生物重组技术的关键是需要使用专门的重组基因载体——质粒。质粒(plasmid)是一种细胞质遗传因子,其具有稳定的遗传特性。相较于动物和植物转基因技术,微生物重组技术具有研发周期短、结果显著以及可控制性强的特点,目前已经广泛应用于生物农药、生物医药、食品发酵以及酶制剂等行业。经过几十年的研究和产业化积累,目前已经开发出大肠杆菌表达系统、酵母表达系统以及丝状真菌表达系统,这些成熟的微生物表达系统已经成功应用于各领域的科研和实际生产中。其中毕赤酵母表达系统和大肠杆菌表达系统最受科研工作者的欢迎,这两个系统其目标产物表达效率高,通常外源目的蛋白占细胞总蛋白的10%~40%,同时这两个系统构建的重组微生物的生产成本低,目前我们常见医药和食品等产品,诸如胰岛素、抗生素、重组人p53腺病毒注射液、啤酒酵母乙肝疫苗、α-高温淀粉酶、饲料用木聚糖酶和壳聚糖酶等产品都由这两种表达系统所生产的(张迎庆等, 1988)。
2转基因的转化技术
按照是否需要通过组织培养的方式获得新生体,转基因的遗传转化方式可分成两大类,第一类方法获取再生植株需要通过组织培养,目前研究中主要方法有基因枪法和农杆菌介导转化法;第二类方法不需要经过组织培养获得新生体,主要有花粉管通道法。
2.1农杆菌介导转化
农杆菌是一种革兰氏阴性细菌,其普遍存在于自然界的土壤环境中的,在自然条件农杆菌能够感染大多数双子叶植物的受伤部位,然后通过一系列反应诱导产生发状根或冠瘿瘤。
根癌农杆菌的细胞中含有Ti质粒,而发根农杆菌含有Ri质粒,在两类质粒上存在一段T-DNA,通常情况下农杆菌通过植物的伤口侵袭进入细胞后,进而将T-DNA插入到靶标植物的基因组中(图5)。
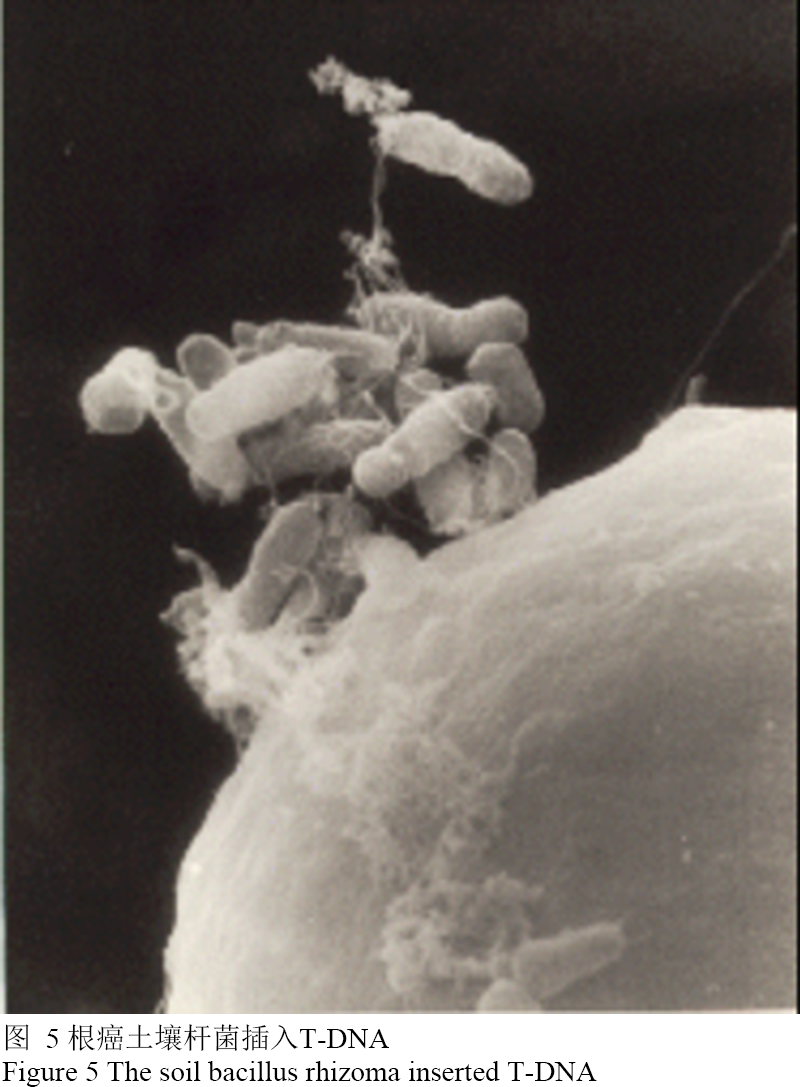 图 5 根癌土壤杆菌插入T-DNA Figure 5 The soil bacillus rhizoma inserted T-DNA |
实际上,农杆菌其本身就是一种天然的植物遗传转化体系,是自然界在进化过程中发展形成的天然“转基因体系”。科学家将目的基因整合入经过改造的T-DNA区,通过农杆菌的感染能力而实现外源基因与靶标植物细胞的整合,然后通过组培等途径,获得再生转基因植株。农杆菌介导法起初只被应用于双子叶植物中,自从农杆菌运用技术瓶颈被打破之后,农杆菌介导转化法从最初的双子叶植物应用扩展到单子叶植物,目前水稻已经被当作科学研究中的模式植物。
2.2花粉管通道法
花粉管通道法于80年代初期由中国学者周光宇提出,其主要的技术途径首先在植物授粉后,通过注射的方式将含目的基因的DNA溶液注射入子房,而外源DNA利用植物在开花和受精过程中形成的花粉管通道而进入受精卵细胞,并通过进一步的整合将外源DNA整合到受体细胞的基因组中,通过生长而形成新的转基因个体(图6)。该方法的的优势在于其不依赖组织培养而获得再生植株,整体技术简单,对于设备的要求较低,是常规育种工作者的首先(Huang et al., 1999)。中国目前种植最为广发的转基因抗虫棉就是通过花粉管通道法培育出来的。
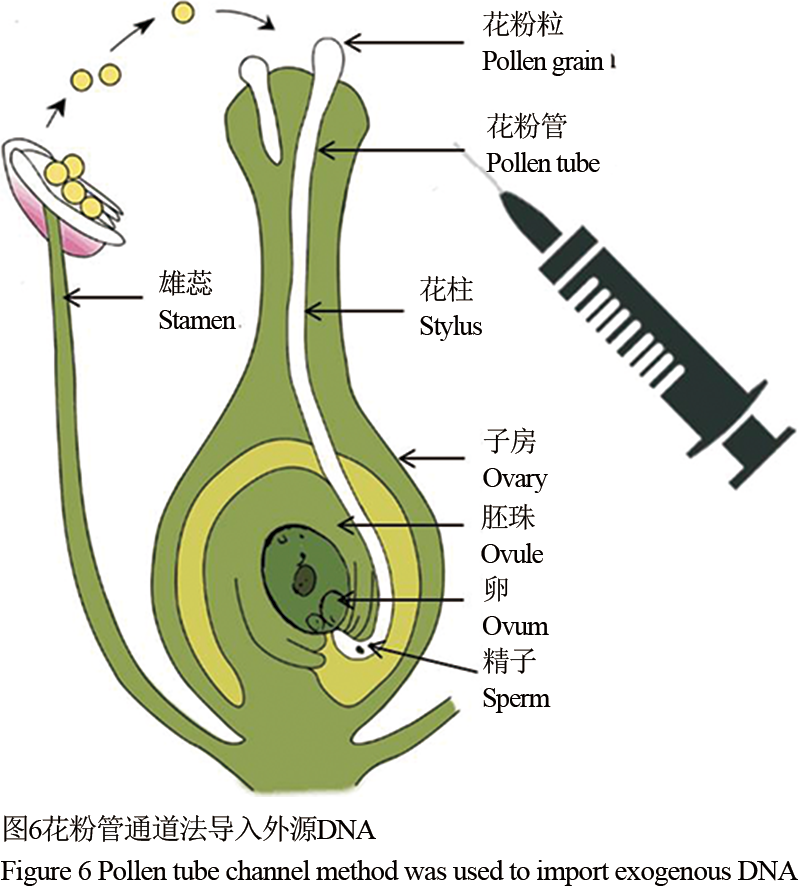 图 6 花粉管通道法导入外源DNA Figure 6 Pollen tube channel method was used to import exogenous DNA |
2.3核显微注射法
核显微注射法是目前动物转基因技术研究中广泛使用的方法,其主要是在显微镜下将外源DNA注射到靶标受精卵细胞的细胞核内,外源DNA整合进入胚胎基因组,通过体外培养后再移植到受体母畜子宫内进行受孕发育。但是这种方法具有效率低、不确定性高、动物利用率低等缺点。用这种方法生产的动物约有十分之一是整合外源基因的转基因动物(图7)。
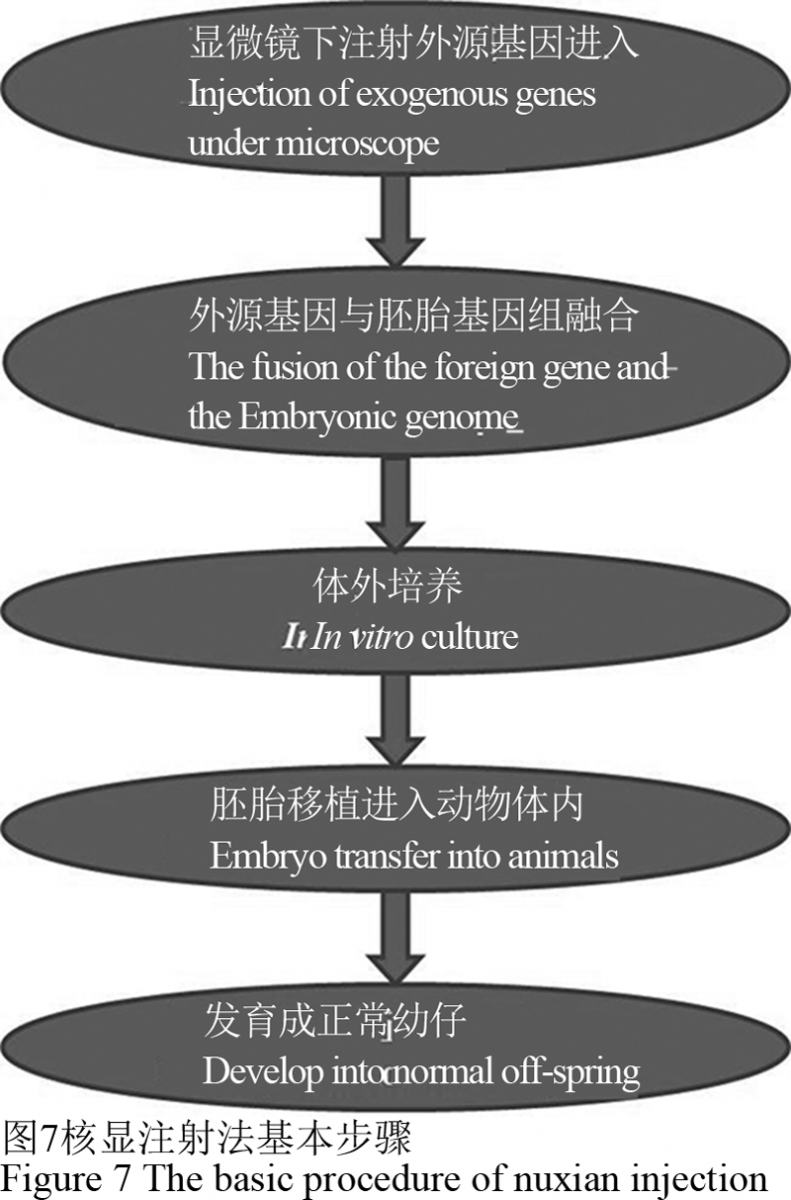 图 7 核显注射法基本步骤 Figure 7 The basic procedure of nuxian injection |
2.4基因枪法
基因枪是指利用火药爆炸或高压气体将带目的基因DNA溶液的高速微弹加速的一种设备,其可以将将外源DNA直接送入完整的植物组织或细胞中,然后利用组培技术获得再生转基因植株。相较于农杆菌转化法,基因枪法转化的一个主要优势在于其不受受体植物种类范围的限制。同时含有外源DNA的载体质粒的构建方法也较为常规和简单,因此该方法是目前转基因研究中应用广泛的一种方法。
基因枪可以在大多数细胞上有效使用,但主要用于植物细胞。基因枪作用机理:首先是基因枪装置准备开火(步骤1);氦气填充腔室,压力建立在破裂盘上(步骤2);当压力最终达到破裂盘破裂的程度,由此产生的氦气爆发推动DNA /镀金大载体('塑料盘')进入停止筛网(步骤3);当大载体撞击停止筛选时,DNA包被的金颗粒被推进筛网并进入靶细胞(步骤4) (图8)。
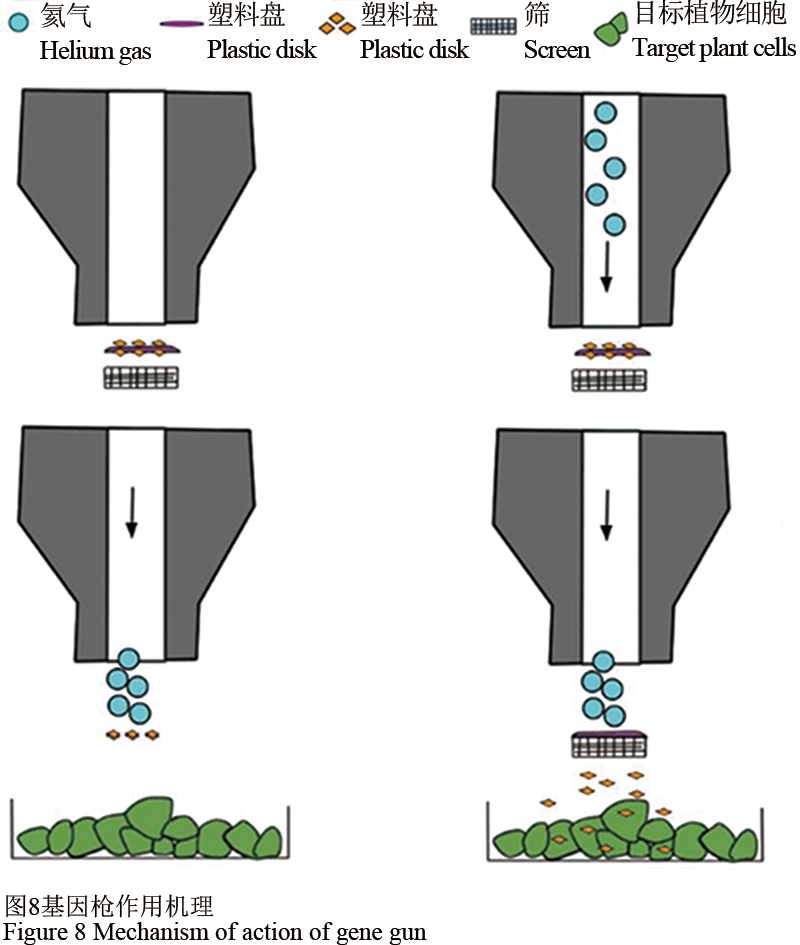 图 8 基因枪作用机理 Figure 8 Mechanism of action of gene gun |
2.5精子介导法
精子介导法是对载体精子进行适当的处理,使该精子具备携带外源基因的能力。然后,利用携带有外源基因的精子受精给发情的母畜。因此,在后代中就会存在一定比例的转基因动物。
相较于显微注射法,精子介导的基因转移存在两个优势:其一是该方法的成本很低,只有显微注射法成本的1/10。其二,该方法并不不涉及对动物进行处理,因此可以对动物群体进行操作,成功率相较更高。
2.6核移植转基因法
体细胞核移植是先把外源基因与供体细胞进行共培养,促使外源基因自然整合到供体细胞的细胞核上,然后将重组后的细胞核移植到受体细胞——去核卵母细胞,构建重组胚,再将重组胚移植到代孕母体,最终可以得到转基因的克隆动物(图9)。
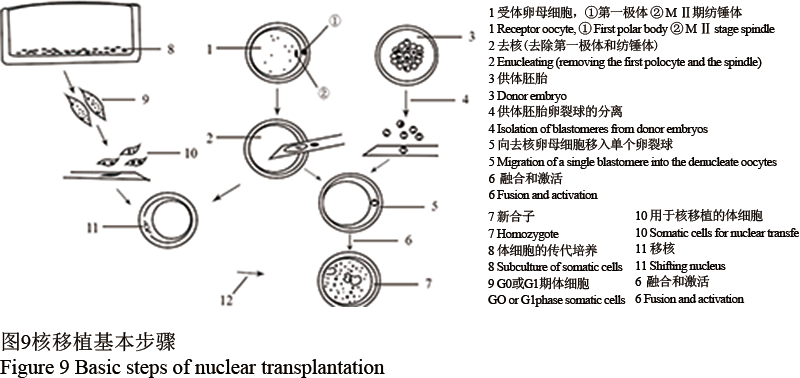 图 9 核移植基本步骤 Figure 9 Basic steps of nuclear transplantation |
2.7体细胞核移植法
体细胞核移植法是先在体外培养的体细胞中导入外源基因,通过筛选进而获得携带目的基因的重组细胞。然后,将带转基因的重组体细胞的细胞核移植到已经去除细胞核的卵细胞中,产生重构胚胎。该种重构胚胎重新移植到受孕母体中,其产生的后代完全是成功的转基因动物,如世界上第一头克隆羊多莉(图10)。
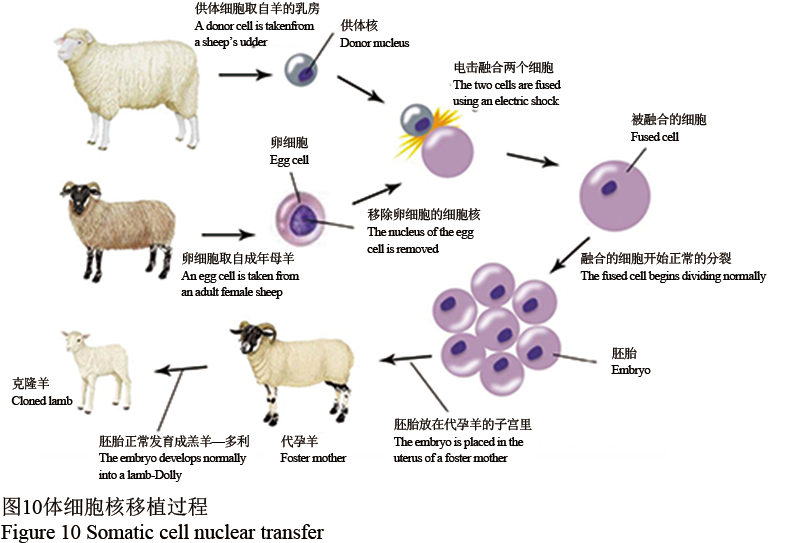 图 10 体细胞核移植过程 Figure 10 Somatic cell nuclear transfer |
2.8基因编辑技术
基因编辑技术是指通过分子生物学途径,人类能够自由的对目的基因进行人为“编辑”,进而实现对特定的DNA片段进行敲除、加入、转移等改变的一种基因操作方法。
基因编辑技术中,以锌指核酸内切酶ZFN (zinc-finger nucleases)和类转录激活因子效应物核酸酶TALEN (transcription activator-like effector nucleases)为代表的序列特异性核酸酶技术以其能够高效率地进行定点基因组编辑,在基因研究、基因治疗和遗传改良等方面展示出了巨大的潜力。
CRISPR/Cas9是继ZFN和TALEN之后出现的第三代“基因组定点编辑技术”。相较于前两代基因编辑技术,CRISPR/Cas9具有成本低、快捷高效和制作简便等优点,已经成为目前基因组研究领域最为火热的科研工具。CRISPR/Cas9技术被认为能够在活细胞中实现最有效、最便捷地“编辑”任何基因(图11)。
.png) 图 11 CRISPR/Cas9技术原理 Figure 11 Principle of CRISPR/Cas9 technology |
3转基因的外源基因
转基因过程中所涉及的外源基因又称为外源DNA,是通过基因工程技术或病毒感染等途径引入靶细胞中的DNA序列。
对于一个细胞来说,内源DNA是其基因组的序列(本身生物就有的DNA),而外源的DNA是通过基因工程导入的其他物种或细胞的DNA,也可以是人工合成的一段DNA。
3.1内参基因
内参基因通常用来作为基因表达内部参照物,这些内参基因在各受体的组织和细胞中的表达相对较为稳定,可以作为参照物来说明目的基因的表达水平,在目前的转基因研究中,不用物种所选用的内参基因不同(表1)。
 表 1 植物转基因研究中常用内参基因 Table 1 Internal reference genes are commonly used in plant transgenic research |
3.2外源调控序列
在DNA中,基因表达的调节通常发生在RNA生物合成(转录)水平,并且通过激活或抑制转录的蛋白质(转录因子)的序列特异性结合来实现。转录因子可以作为激活剂,抑制剂或两者。抑制剂通常通过阻止RNA聚合酶与转录起始区(启动子)形成生产性复合物起作用),而激活剂促进了生产性复合物的形成。此外,已显示DNA基序可预测表观基因组修饰,表明转录因子在调节表观基因组中起作用。外源调控序列指通过人为导入,具有调控基因表达功能的一段DNA序列,常见的有各种启动子、终止子、TATA框(TATA box)、CAAT框(CAAT box)、增强子等元件(表2)。
 表 2 转基因研究中常见外源调控序列 Table 2 Exogenous regulatory sequences are common in transgenic studies |
3.3外源目标基因
靶基因即目标基因,外源目标基因即来源自外部而能对本体发生作用的目标基因。在分子遗传中,外源目标基因的导入,主要为实现改变生物特性的目标,是主要的性状变化基因。根据研究者的内容需求不同,所选择的外源目标基因的种类也不同(表3)。
 表 3 转基因研究中常见外源目标基因 Table 3 Common foreign target genes in transgenic research |
4转基因的应用
目前,转基因技术已经广泛应用于农业、工业、医药、环保、能源和新材料等众多领域(McAfee, 2003)。
4.1药物领域
目前通过基因工程技术,已有创造出基因工程胰岛素、基因工程疫苗以及基因工程干扰素等药物(Vane and Cuatrecasas, 1984)。科学家们通过转基因技术,人为定向改造植物、动物或者微生物的遗传性状,将其改造成一个“生物反应器”,进而生产人类所需要的基因产物,即药物原料和药品(Powell and Buchanan, 2011)。
自从1982年世界上第一个转基因工程药物——重组人胰岛素,经美国FDA批准上市以来(Ferrer-Miralles et al., 2009),基因工程药物得到了飞速发展。
4.1.1基因工程疫苗
使用DNA重组生物技术,把人工合成或者来自其他物种的天然遗传物质定向整合进入细菌、酵母菌或哺乳动物的基因组中,使外源基因得到充分表达,获取表达产物经过纯化后而制得疫苗,称为基因工程疫苗。通过应用基因工程技术,可以按照人类的设想意愿,研制出不含感染性物质的亚单位疫苗、脱毒疫苗及能够对多种疾病防疫的多价疫苗。目前乙肝疫苗(Szmuness et al., 1981)、百日咳基因工程疫苗、狂犬病基因工程灭活疫苗(Ito et al., 2005)、产肠毒素大肠杆菌基因工程疫苗、Asia Ⅰ型口蹄疫病毒(FMDV)的感染表位重组蛋白疫苗(Mackay et al., 1988)以及肠出血性大肠杆菌基因工程疫苗等已经成为广发使用的商业化疫苗。
4.1.2基因工程胰岛素
截止到2013年,中国的糖尿病患者人数达1.14亿,占全球糖尿病患者的1/3。糖尿病的致病原因就是胰岛素分泌缺陷,或其他太代谢途径受损,目前临床上最为常用的治疗方法就是以人工注射胰岛素的途径从体外补充患者所需的胰岛素。胰岛素最初只能从哺乳动物牛和猪的胰脏中提取,但是产量非常低,每100 kg牛或者猪的胰腺只能提取出4~5 g的胰岛素,这样的产量远远不能满足患者的需求。
1980年代初,美国一家公司利用转基因技术,在微生物体内实现了人体胰岛素的工业化生产(Keen et al., 1980)。该研究团队通过克隆技术获取人基因组中表达胰岛素的那一段DNA,将该段DNA人工转入大肠杆菌或者酵母菌里,构建重组表达工程菌,利用微生物快速繁殖的特性,进而获得大量的人体胰岛素,极大的降低了胰岛素的生产成本,造福了全球的糖尿病患者。
此外,基因工程乙肝疫苗也通过转基因技术而实现了产业化生产。中国卫计委在2013年7月26日公布,全球目前存在3.5亿乙肝病毒携带者,其中中国有近1亿携带者;在全球每年大约70万因病毒性肝炎而致死的患者中中国占近半数。20世纪80年代,研究者将乙肝病毒基因中表达表面抗原的那一段DNA“剪切”成功克隆,并转入酵母菌里或者重组表达菌。当酵母菌正常生长时,被转入的乙肝病毒基因就会随之而生产出乙肝表面抗原,并随着酵母菌的快速大量繁殖而量产。这种疫苗技术于1994年被引进中国,随后建成了两条基因工程乙肝疫苗生产线,在2001年以后,中国医药领域使用的全部是高安全性的基因工程乙肝疫苗。中国政府也开始着手给全国的儿童免费接种、甚至免费补种乙肝疫苗。根据中国卫计委的数据统计显示,1992年至2009年中国预防了近8000万人免受乙肝病毒感染,减少了近2000万乙肝病毒表面抗原携带者,减少肝硬化、肝癌等引起的死亡430万人。
4.1.3食品领域
利用现代分子生物学技术,人为设计的将某些生物的具有特殊性状的基因转移到另一种农作物中去,通过基因重组而定向改造生物的遗传性状,使转基因农作物在性状和营养品质等方面转变为人类所需要的。转基因食品在广义上可以囊括以转基因生物为直接食品,或以转基因生物作为原料而加工产生的食品,以及用转基因生物喂养家畜得到的衍生食品。但就目前来看,历经几十年的发展,转基因食品因其安全性未能够获得定论,在国际社会上仍存在广泛质疑和争论。
尽管转基因食品的科学研究已有几十年的历史,但是转基因成功的商业化才是近二十年的事情。20世纪90年代初,在美国的市场上出现了第一个转基因食品——保鲜番茄,展示出了非常强劲的市场效应,此后,转基因食品迅速发展,逐渐成为食品领域的新宠儿。据统计,美国食品和药物管理局登记批准进入市场的转基因品种已达到43种。
如常见的Bt (苏云金芽孢杆菌, Bacillus thuringiensis)转基因抗虫农作物和转Ht基因抗除草剂作物,已经成为转基因抗性农作物的代表。Bt菌株能够分泌产生一种对鳞翅目、鞘翅目等农业害虫具有高效杀虫活性的毒素蛋白质,这种毒蛋白只对昆虫有效,目前尚未有充足的证据能够证明其对人类或其他哺乳动物产生致毒或者致敏的毒性。目前,转Bt基因抗虫作物如抗虫棉、抗虫玉米和抗虫大豆等在全球获得广泛的种植。Ht基因是一种抗除草剂基因,该基因编码的蛋白质能够分解植物体内的除草剂药物,使植物获得抵抗高浓度除草剂的能力,免受除草剂农药的伤害。因此在大田喷洒除草剂之后,没有除草剂抗性的杂草会因为对除草剂的抵抗力不足而被农药杀死,而转Ht基因的农作物因为自身的抗性而得以正常存活。相对于传统的机械性除草,种植转Ht基因的农作物能够减少人工的投入而更加经济。
截至2013年,对于转基因食品的分类尚没有明确的标准,存在多种不同的分类标准。
根据转基因食品中是否含有转基因源为标准可分为如下3种不同类型:
(1) 食品本身不含转基因的转基因食品,是指食品的来源是转基因生物,但该食品本身并没有携带任何外源基因。
(2) 转基因食品中含有非活性的转基因成分,是指食品来源存在转基因,但是在该食品加工过程中转基因的特性已发生了改变,不存在于人类食用或食用的食品中。
(3) 转基因食品中带有活性的基因成分,是指人们食用的这种转基因食品,其本身携带有具有活性的转基因产物,而且均会被人体消化吸收。
根据转基因食品来源的不同可分为如下三种不同类型:
(1) 植物性转基因食品,是指以含有转基因成分的植物作为原料的直接或加工后的转基因食品。
(2) 动物性转基因食品,是指以含有转基因成分的动物作为原料的直接或加工后的转基因食品。
(3) 微生物转基因食品,是指以含有转基因的微生物为原料的转基因食品。
根据食品中转基因的功能的不同大致可以分成以下6种类型:
(1) 增产型的转基因食品
(2) 控熟型的转基因食品
(3) 保健型的转基因食品
(4) 加工型的转基因食品
(5) 高营养型的转基因食品
(6) 新品种型的转基因食品
5关于转基因的若干概念
随着分子生物技术的不断发展,尤其是上世纪90年代末以来,科学家们能够利用“基因组定点编辑技术”在不导入外源基因的情况下,仅仅通过对目标生物体本身的遗传物质进行加工、敲除和屏蔽等方法途径,也能人为有效的改变生物体的遗传特性,从而获得研究者们所希望得到的性状。然而,这与传统的转基因定义产生了一定的冲突,通常来说没有转入外源基因,就不能称之为为转基因,而称为“基因修饰”似乎会更加合适和全面,因此现在开始用“Genetically Modified Organisms”(简称GMOs),即“基因修饰生物”,来代替早期的“Transgenic Organisms”。因此,现在我们所指的“转基因生物”,其概念已经为“基因修饰生物”所涵盖。但因为“转基因”一词历经几十年的发展,已经普遍为人们所接受,而且通过外源基因导入改变生物体性状的方式,仍然是目前基因工程作物育种领域中所采用的主要方法之一,所以“转基因生物”一词就沿用至今。基于此,本综述继续沿用“转基因”一词,不过是在“基因修饰生物”的意义上加以阐述和使用。
6结束语
转基因是现代科学发展的必然,当人类掌握操纵基因的技术后,必定会通过转基因技术,为自身发展谋福利,如增强营养、提高产量、减少农药用量等。另一方面,相比日新月异的现代科技,普通民众的知识增长相对落后,且易受不可知论的蒙蔽,加之新技术对传统利益集团的冲击,反转基因运动的兴起也是自然而然的事情。所有涉及转基因研究的群体,必须要以对人类高度负责的态度来积极研究、审慎应用、严格管理,推进转基因健康发展,把科学技术更好地造福人类,增进人类的福祉。
作者贡献
傅学彪负责文献收集、稿件修改和补充;吴仲琦进行论文构思和初稿写作、修改和最终定稿。全体作者都阅读并同意最终的文本。
致谢
本研究诸暨市翠溪生物技术研究院科普项目基金资助。
Arias, D. M., and Rieseberg, L. H., 1994, Gene flow between cultivated and wild sunflowers, Theoretical and Applied genetics, 89(6): 655-660
https://doi.org/10.1007/BF00223700
Berge, J., Gabrielsen, T. M., Moline, M., & Renaud, P. E., 2012, Evolution of the Arctic Calanus complex: an Arctic marine avocado?, Journal of plankton research, 34(3): 191-195.
https://doi.org/10.1093/plankt/fbr103
Chang A C Y., and Cohen S N., 1974, Genome construction between bacterial species in vitro: replication and expression of Staphylococcus plasmid genes in Escheric hia coli, Proceedings of the National Academy of Sciences, 71(4): 1030-1034
https://doi.org/10.1073/pnas.71.4.1030
Ferrer-Miralles, N., Domingo-Espín, J., Corchero, J. L., Vázquez, E., and Villaverde, A., 2009, Microbial factories for recombinant pharmaceuticals, Microbial cell factories., 8(1): 17
https://doi.org/10.1186/1475-2859-8-17
Hinnen A., Hicks J B., and Fink G R., 1978, Transformation of yeast, Proceedings of the National Academy of Sciences, 75(4): 1929-1933
https://doi.org/10.1073/pnas.75.4.1929
Huang, G., Dong, Y., and Sun, J., 1999, Introduction of exogenous DNA into cotton via the pollen-tube pathway with GFP as a reporter, Chinese science bulletin, 44(8): 698
https://doi.org/10.1007/BF02909705
Ito, N., Sugiyama, M., Yamada, K., Shimizu, K., Takayama‐Ito, M., Hosokawa, J., and Minamoto, N., 2005, Characterization of M gene‐deficient rabies virus with advantages of effective immunization and safety as a vaccine strain, Microbiology and immunology., 49(11): 971-979
https://doi.org/10.1111/j.1348-0421.2005.tb03692.x
Keen, H., Pickup, J. C., Bilous, R. W., Glynne, A., Viberti, G. C., Jarrett, R. J., and Marsden, R., 1980, Human insulin produced by recombinant DNA technology: safety and hypoglycaemic potency in healthy men, The Lancet, 316(8191): 398-401
https://doi.org/10.1016/S0140-6736(80)90443-2
Ledford, H., 2016, CRISPR: gene editing is just the beginning, Nature News., 531(7593): 156
https://doi.org/10.1038/531156a
Mackay, D. K. J., Forsyth, M. A., Davies, P. R., Berlinzani, A., Belsham, G. J., Flint, M., and Ryan, M. D., 1998, Differentiating infection from vaccination in foot-and-mouth disease using a panel of recombinant, non-structural proteins in ELISA, Vaccine., 16(5): 446-459
https://doi.org/10.1016/S0264-410X(97)00227-2
McAfee, K., 2003, Neoliberalism on the molecular scale, Economic and genetic reductionism in biotechnology battles. Geoforum., 34(2): 203-219
https://doi.org/10.1016/S0016-7185(02)00089-1
Powell, R., and Buchanan, A., 2011, Breaking evolution's chains: the prospect of deliberate genetic modification in humans, In The Journal of Medicine and Philosophy: A Forum for Bioethics and Philosophy of Medicine., 36(1): 6-27
https://doi.org/10.1093/jmp/jhq057
Shou, H., Palmer, R. G., and Wang, K., 2002, Irreproducibility of the soybean pollen-tube pathway transformation procedure, Plant molecular biology reporter., 20(4): 325-334
https://doi.org/10.1007/BF02772120
Szmuness, W., Stevens, C. E., Zang, E. A., Harley, E. J., and Kellner, A., 1981, A controlled clinical trial of the efficacy of the hepatitis B vaccine (Heptavax B): a final report, Hepatology., 1(5): 377-385
https://doi.org/10.1002/hep.1840010502
Vane, J., and Cuatrecasas, P., 1984, Genetic engineering and pharmaceuticals, Nature., 312(5992): 303
https://doi.org/10.1038/312303a0
Zhang Y.Q., Gan X., Fang S.L., and Yu D.X., 1988, Application of Molecular Cloning Technique in Antibiotic Engineering Strains, Journal of Hubei Polytechnic University, 13(04): 33-37 (张迎庆, 干信, 方尚玲, 余东霞, 1988, 分子克隆技术在抗生素工程菌中的应用, 湖北工学院学报, 13(04): 33-37)
Zhou X.P., 2011, Study of HBV DNA integration in oocyte/embryo and the ART outcomes of HBV infected women, Zhejiang university (周旭平, 2011, 乙型肝炎病毒DNA在人卵母细胞和胚胎中的整合及HBV感染妇女ART结局的研究, 浙江大学)