研究报告/Research Report
基于RNA-Seq数据筛选的小桐子抗冷相关基因的表达模式及其聚类分析 
2曲靖师范学院生物资源与环境科学学院, 曲靖, 655011
 作者
作者  通讯作者
通讯作者
植物药与药理学杂志, 2014 年, 第 3 卷, 第 2 篇
收稿日期: 2014年12月05日 接受日期: 2014年12月16日 发表日期: 2014年12月28日
能源树种小桐子属冷敏植物,受低温胁迫影响涉及到形态、生理、生化、基因表达及调控等多个方面。我们最近通过RNA-Seq技术获得了小桐子低温锻炼转录组(45 251条Unigene)及其数字基因表达谱数据,发现在12℃低温锻炼下有4 185个基因表达量发生变化,其中在不同锻炼时间(12 h, 24 h, 48 h)内都发生变化的基因有553个。本工作从中选择了22个与对照相比有较大表达差异、新诱导的或被公认的抗冷相关基因,通过半定量RT-PCR及进一步聚类分析检测它们在小桐子不同组织及低温锻炼过程中的表达模式,结合其数字基因表达结果以期找出与小桐子抗冷性密切相关的潜在关键基因,为小桐子抗冷性的遗传改良打下基础。
起源于中南美洲热带地区的油料树种小桐子在世界范围内都是重要的能源植物(Lin et al., 2004; 何璐等, 2010)。但是,由于其热带习性,小桐子耐寒性差,低温已成为限制小桐子生长和种植分布的主要环境因子,也是制约小桐子产业发展的瓶颈因素(曾觉民, 2006)。通过各种生理或分子育种技术培育抗冷新品种预防冷害并扩大小桐子在高纬度低温气候环境下的种植面积,已成为目前亟待解决的问题,因此深入研究小桐子的抗冷分子机制具有重要的理论与现实意义。
我们先前研究结果表明:低温锻炼能显著降低小桐子幼苗死亡率、叶片电解质渗透率以及MDA的含量,有效减轻冷胁迫对膜系统的伤害和降低膜脂过氧化损伤,而且12℃低温锻炼48h对提高小桐子幼苗抗冷性的效果最佳;同时,低温锻炼能显著提高小桐子抵御ROS伤害的能力,伴随着抗氧化酶(SOD, CAT, APX, GR, POD, DHAR, MDAR)活性、抗氧化物质(AsA, GSH)浓度、AsA/(AsA+DHA)和GSH/(GSH+GSSG)比值的提高(Ao et al., 2013a; 2013b);这些与Foyer 和 Noctor (2009)的研究相一致。另外,我们最近通过RNA-Seq技术获得了小桐子低温锻炼转录组与数字基因表达谱的测序与分析结果(Wang et al., 2013; 2014)。为了准确深入阐明低温锻炼在提高小桐子抗冷性中的分子机制,本工作从中筛选了22个低温锻炼与对照相比差异表达较大的基因、新诱导表达的基因以及一些公认的与低温响应密切的基因,通过半定量RT-PCR调查了这些基因在小桐子不同组织和低温锻炼时间内的表达模式并进行聚类分析,试图结合基因的表达模式变化与其数字基因表达谱的结果,找出与小桐子抗冷性可能密切相关的关键基因,为后续的小桐子抗冷性遗传改良研究奠定基础。
1结果与分析
1.1基于RNA-Seq数据筛选小桐子抗冷相关的代表基因
本实验室最近报道了通过RNA-Seq技术获得比较完整的小桐子低温锻炼转录组(45 251条Unigene)及其数字基因表达谱数据(Wang et al., 2013; 2014),发现在12℃低温锻炼下有4 185个基因呈现差异表达,其中在不同锻炼时间(12h, 24h, 48h)内都发生表达变化的基因有553个。为了准确深入阐明低温锻炼在提高小桐子抗冷性中的分子机制,本工作特地从中筛选了22个与桐子抗冷相关的代表基因(表1),包括与对照相比差异表达较大基因(Ⅰ类)、新诱导表达基因(Ⅱ类)、公认的抗冷相关功能基因(Ⅲ, Ⅳ, Ⅴ, Ⅵ类)以及抗冷相关转录因子基因(Ⅶ类),并通过GenBank进行BLAST功能注释验证。根据它们的Unigene序列设计特异引物(表2),并以18S rRNA为内参通过半定量RT-PCR进行不同组织和不同低温锻炼时间内的表达模式分析。
|
表 1 表达模式研究涉及的小桐子抗冷相关基因 |
.png)
|
表 2 引物序列信息 |
.png)
1.2小桐子抗冷相关基因在正常条件下不同组织中的表达特性
以18S rRNA为内参,采用半定量RT-PCR分析了候选的22个抗冷相关基因在正常生长小桐子不同组织(根、茎、叶)中的表达情况(图1);通过Quantity ONE软件对其做定量分析,并根据内参值进行校准得到其相对表达量(数据未显示)。结果表明,候选的基因存在明显的组织表达特异性,大部分在茎中表达量较高,而在叶中表达量较低,甚至部分基因没有表达。在根中,22个基因中有16个明显表达,尤其是nsLTPA、POD73、DNAj、GA20ox、ADMT、DXS、FAD3、PDI、LEA5、NAC、WRKY的表达量达到了较高水平;在茎中,22个基因有20个明显表达,尤其是nsLTPA、POD73、UGT、DNAj、GA20ox、ADMT、DXS、Ca2+-ATPase12、FAD3、PIP2、NR、LEA5、NAC、WRKY的表达量达到了较高水平;在叶中,各基因的表达量相对较低,且有很大部分基因(如Cyt P450, UGT, DNAj, ADMT, DXS)没有表达,22个基因中仅有10个基因明显表达,达到高表达的有nsLTPA、GSTs、FAD3、PIP2、NR、PDI、LEA5、NAC、WRKY。综合看来,在3种组织中都有表达的基因包括nsLTPA、POD73、FAD3、PIP2、DHN2、LEA5、NAC、WRKY,说明这8个基因在小桐子中属于组成型表达。
|
图 1 小桐子抗冷相关基因在正常条件下不同组织中的表达 |
.png)
从基因功能角度上分析,候选的22个抗冷相关基因中,在根中表达量最高的5个基因分别为FAD3、LEA5、POD73、DNAj、NAC,前三者分别属于脂肪酸去饱和增加膜流动性相关基因、增加植物细胞吸水能力的基因、抗氧化保护酶基因,都与植物抗冷性直接相关;后两个分别属于分子伴侣蛋白基因与转录因子基因。在茎中,表达量最高的5个基因分别为NR、POD73、LEA5、NAC、GA20ox,其中NR、GA20ox也属于植物抗氧化保护酶系统。在叶中,表达量最高的5个基因分别为PIP2、PDI、NR、LEA5、FAD3,其中PIP2属于促进植物吸水能力的基因,PDI则属于植物抗氧化保护酶系统。在三种组织中组成型表达的8个基因可以分别归类为抗氧化系统保护酶基因(如POD73, DHN2)、植物促进物质运输尤其是水与脂类运输的基因(如nsLTPA, PIP2, LEA5)、以及与信号转导系统相关的转录因子基因(如WRKY)。
1.3小桐子抗冷相关基因在不同组织和低温锻炼时间点的表达模式分析
我们接着采用半定量RT-PCR,以18S rRNA为内参,分析了22个小桐子候选抗冷相关基因在不同低温锻炼时间点(12 h, 24h, 48 h)于不同组织(根, 茎, 叶)中的表达情况(图2),并通过Quantity ONE软件定量和根据内参值校准计算其相对表达量(数据未显示),所得结果如下:
|
图 2 小桐子抗冷相关基因在不同组织和低温锻炼时间点的表达模式 |
.png)
在筛选的低温锻炼与对照相比差异表达较大的基因(Ⅰ类)中,各基因在叶中的表达差异性最明显;而在根、茎中各基因的表达量普遍较叶中高,其中nsLTPA、POD73、UGT、GSTs在茎中组成型表达,随着低温锻炼时间的延长表达量呈明显上调,在24 h达到最大值,但到48 h时又逐渐下降、不过仍高于对照。在根中各基因的表达水平介于茎与叶之间,其中CytP450、UGT、GSTs的表达量随着低温锻炼时间的延长持续增加,而POD73、nsLTPA的表达与对照相比并没有太大变化,甚至在低温锻炼12 h与48 h时还略有下调。
在筛选的低温新诱导表达基因(Ⅱ类)中,各基因在叶中于对照条件下都几乎没有表达,而随着低温锻炼时间的延长逐渐增加,到24 h时大部分基因达到表达最高值,随后部分基因(如DNAj, GA20ox)持续高表达,而部分基因(如CHS, LRR-RLK, ADMT, DXS)表达量则逐渐下降。此类基因在茎中表达差异性很大,但在低温锻炼的不同时间点各基因的表达量与对照基本持平,个别基因(如ADMT, DXS)还有下调趋势。与茎、叶中表达相比,该类基因在根中差异表达也较明显,多数基因在低温锻炼条件下,与对照相比表现出先下调再上调的变化趋势,如DNAj、GA20ox、CHS、ADMT、DXS,而LRR-RLK基因在低温锻炼48 h时才明显上调。
FAD3 (Ⅲ类)是催化植物体内由二烯酸生成三烯酸的主要酶之一,受低温诱导表达。半定量RT-PCR结果表明,FAD3基因在对照条件下都有本底表达;在低温锻炼12 h时,根、茎及叶中表达量都明显下调;但随着低温锻炼时间的延长,其表达量又逐渐上升,表现出对低温的响应。APX1 (Ⅳ类)与PIP2 (Ⅴ类)分别是与低温应答相关的抗氧化酶系统与水分运输膜蛋白家族的关键基因。与对照相比,在根中,随着低温锻炼时间的延长,其表达量与先下调再上调,到24 h时达到最大值;而在茎中,其表达量变化较小,只是APX1在低温锻炼24 h才有明显上调;在叶中,APX1表达量最低,PIP2则表现出随着低温锻炼时间延长而逐渐下调的变化趋势。
另外,在茎中,DHN2、NR、HSP17.5、LEA5基因(Ⅵ类)在低温锻炼下与对照相比的表达量变化不明显,基本上算是持续本底表达,而且PDI基因在茎中基本没有表达。在根中,该类基因于低温锻炼下表现出先下调后上调的表达变化趋势,如DHN2、NR、LEA5,在低温锻炼12 h时进入低温适应阶段,而随着低温时间的延长其表达量逐渐上调,以抵抗低温对植物的进一步伤害。而在对低温更加敏感的叶片中,NR、HSP17.5表现出低温锻炼瞬间诱发表达增加现象,且随着低温时间延长而持续表达。在这类基因中,PDI明显表现出组织表达特异性,在茎中基本无表达,在根中受低温诱导表达,而在叶中保持组成型表达;而DHN2在叶中表达量很少,而在根、茎中又受到低温诱导表达。这些结果充分反映出不同基因在不同低温锻炼条件下在不同组织中的表达复杂性。
NAC与WRKY转录因子(Ⅶ类)是除ICE、CBF之外、与植物抗冷性最密切的转录因子,都参与植物依赖ABA的冷信号转导途径,是接收低温初始信号并向下游传递的关键调节子。半定量RT-PCR结果表明,与对照相比,NAC、WRKY在茎中于不同低温锻炼时间下的表达量基本保持本底水平;而在根与叶中的表达变化基本一致:即随着低温锻炼时间的延长表现出先下调后上调的变化趋势,尤其是NAC在低温锻炼24 h时的表达量上调达到了极显著水平。
1.4小桐子抗冷相关基因表达模式的聚类分析
分为更好展示小桐子抗冷相关基因在低温锻炼过程中的表达变化规律,我们使用K-mean算法对这些候选基因进行了聚类分析。根据上述1.3中基因在低温锻炼过程中差异表达倍数的变化情况,将基因在根、茎及叶中表达变化模式分别聚类成不同类别,(图3)。
|
图 3 小桐子抗冷相关基因差异表达模式的聚类分析 |
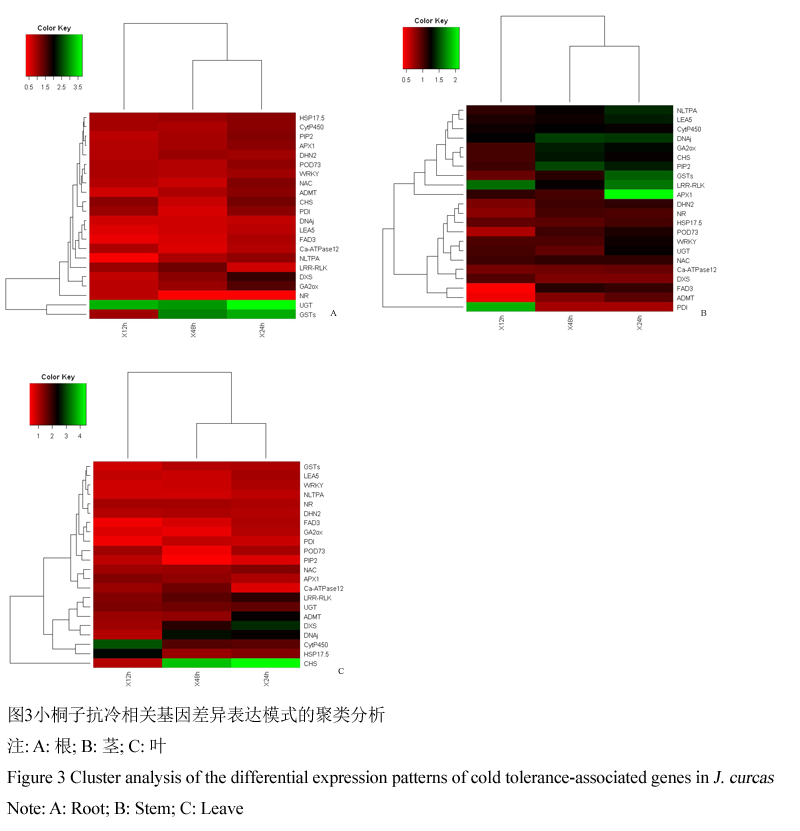
在根中,22个基因按变化趋势聚类为5类(图3A):第一类基因表达量表现为先微降再骤然上升后下降的模式,包括nsLTPA、POD73、DNAj、GA20ox、ADMT、DXS、FAD3、APX1、PIP2、HSP17.5、NAC、WRKY、LEA5 (13个);第二类基因表达量表现为骤升再降低的模式,包括UGT、CHS、PDI、GST (4个);第三类基因表达量表现为先骤升再降低后上升的变化模式,包括CytP450、LRR-RLK (2个);第四类基因表达量表现为先降低后骤然上升的变化模式,包括DHN2、NR (2个);第五类基因表达量表现为持续下降的模式,包括Ca-ATPase。
在茎中,22个基因按变化趋势也聚类为5类(图3B):第一类基因表达量表现为先微降再骤然上升后下降的模式,包括CytP450、POD73、GSTs、ADMT、Ca-ATPase、DHN2、HSP17.5 (7个);第二类基因表达量表现为骤升再降低的模式,包括nsLTPA、UGT、LRR-RLK、APX1、WRKY、LEA5、PDI (7个);第三类基因表达量表现为持续上升的模式,包括DNAj、GA20ox、CHS、PIP2、NAC (5个);第四类基因表达量表现为先降低后骤然上升的变化模式,包括FAD3、NR (2个);第五类基因表达量表现为持续下降的模式,包括DXS。
在叶中,22个基因按变化趋势可聚类为6类(图3C):第一类基因表达量表现为骤升再降低的模式,包括POD73、UGT、LRR-RLK、ADMT、DXS、HSP17.5、NAC (7个);第二类基因表达量表现为先微降再骤然上升后下降的模式,包括nsLTPA、GSTs、GA20ox、FAD3、WRKY、LEA5 (6个);第三类基因表达量表现为先骤升再降低后上升的变化模式,包括APX1、DHN2、NR、Ca-ATPase (4个);第四类基因表达量表现为持续上升的模式,包括CytP450;DNAj、CHS (3个);第五类基因表达量表现为先降低后骤然上升的变化模式,包括PDI (1个);第六类基因表达量表现为持续下降的模式,包括PIP2 (1个)。
2讨论
使用基因表达的变化是调控细胞活动的核心机制,正是由于基因的差异表达才决定了生物体的所有生命过程,包括对外界环境的适应。因此,研究生物体不同类型组织或同一类组织在不同发育时期或不同环境条件下基因表达的变化,已成为分子生物学的主要研究方向之一。与传统的Northern blot法比较,半定量RT-PCR具有灵敏、简捷、廉价和特异性高等优点,在检测基因表达水平上得到了广泛应用,因而用作本工作中小桐子抗冷相关基因表达模式的主要分析手段。同时,本工作进一步采用K-mean算法进行聚类分析,可以更好地展示小桐子抗冷相关基因在低温锻炼过程中的表达变化规律。
在筛选的5个低温锻炼与对照差异表达变化较大的基因(Ⅰ类)中,总体表现为叶中差异表达最明显,在根中表现次之,而在茎中表现最差。这5个基因在各组织中于低温锻炼下表达量都有不同程度的上调,但与小桐子抗冷性相对最关联的基因是UGT与GSTs (图2; 图3)。UGT基因编码UDP-葡萄糖苷转移酶,催化活化的葡萄糖分子(UDPG, 尿苷二磷酸葡萄糖)转移到蛋白质、脂类、寡糖等受体分子上形成糖蛋白、糖脂、低聚糖。在植物抵御低温胁迫过程中,各种低聚糖、寡糖作为渗透调节分子及信号转导分子起重要的中介作用。另外,多数情况下糖基化还是植物合成黄酮类、皂苷类及氰醇等次生代谢物质的最后一步反应(Von Rad et al., 2001),这些次生代谢物质对于植物抵御和适应环境变化起重要作用(Lorenc-Kukula et al., 2005)。UGTs属多基因家族,已知拟南芥中至少有120个成员,水稻中有193个成员,苜蓿中有165个成员(Ko et al., 2006)。GSTs编码谷胱甘肽-S-转移酶,是植物体抗氧化反应以及结合有毒物质与活性氧分子的主要清除蛋白。在烟草中过量表达内源性τ型GSTs (Nt107)后,植株内的GST和谷胱甘肽过氧化物酶(GPX)活性显著提高,转基因烟草对高温、低温、高盐、除草剂等的耐受性也得到了增强(Roxas et al., 2000)。
根据数字基因表达谱结果,我们还选择了7个低温锻炼与对照相比新诱导表达的基因(Ⅱ类)。作为新诱导基因,应该是与小桐子抗冷性形成密切相关的核心基因。在RT-PCR实验中(图2),这7个基因在叶中的表达变化趋势与数字基因表达谱数据基本吻合。这几个基因在对照条件下几乎都没有表达,但随着低温锻炼时间的延长各基因的表达量表现为先缓慢上调,在低温锻炼24 h时表达量都同时达到了最高值,随后又缓慢下调。其中,CHS基因是在叶中于低温锻炼条件下表达量上调最大的基因,并且在根、茎中也有明显上调。在叶中,CHS表达表现为后骤然上升的趋势:在低温锻炼12 h时其表达量与对照基本相同,而在24 h、48 h时其表达量较对照分别上调4.435 3、3.930倍;在茎中,它表现为持续上调表达的趋势:与对照相比,在低温锻炼12 h、24 h、48 h下其表达量分别上调1.034 5、1.239 2、1.325 4倍;而在根中,在低温锻炼24 h时它表达量达到最高值,较对照上调了1.257 7倍(图2; 图3)。这些结果充分表明CHS基因在小桐子抗冷性形成中的关键作用,与Ferrer等(1999)的报导相一致。
另外,我们还结合文献筛选了与植物抗冷性相关的8个功能基因(Ⅲ~Ⅵ类)。决定高等植物抗冷性强弱的一个重要因子是三烯脂肪酸(TA)即C18:3与C16:3的含量,而催化三烯脂肪酸合成的关键ω-3去饱和酶FAD3、FAD7、FAD8的研究已逐渐开展。FAD7表达不受低温的影响;而FAD3与FAD8则对低温敏感,在低温胁迫下受诱导表达(Kodama et al., 2000; Yu et al., 2009)。RT-PCR分析表明,在小桐子中,FAD3基因在根、茎及叶中均表现为先下调后上调的表达模式,且随着低温锻炼时间的延长其表达量基本恢复到对照的表达水平(图2)。PIP是目前在其它植物物种中研究较多的抗逆性相关基因(Lumb and Bulleid, 2002),编码水通道蛋白。研究表明,PIP家族蛋白在植物低温胁迫条件下一般都会上调表达,以增加植物从外界向细胞内的水分运输能力从而提高细胞的保水能力。而在本研究中,小桐子PIP2基因在叶中随着低温锻炼持续下调表达,推测 PIP2可能参与植物细胞中水分的胞外运输。在低温胁迫条件下,小桐子叶片细胞为了保持胞内的水分,通过降低PIP2的表达以减少水分向细胞外的运输,从而间接增加胞内的含水量。
目前,关于植物低温信号转导途径研究最为清楚的是不依赖ABA的途径。结合数字基因表达谱数据,我们选择了两个与植物不依赖ABA途径和依赖ABA途径都相关、而且在抗冷性方面研究较少的转录因子NAC、WRKY (Ⅶ类),以研究它们在低温信号转导系统和小桐子抗冷性形成中的作用。NAC转录因子数量众多, 为植物特有且广泛分布于陆生植物中,构成了最大的转录因子家族之一。目前已发现多个NAC因子参与植物对逆境(如低温, 高温, 干旱等)的响应(Zheng et al., 2009)。在本研究中,小桐子NAC转录因子在根、茎及叶中于低温锻炼下都表现为上调表达(图2; 图3),尤其在叶中12℃低温锻炼12 h、24 h、48 h下其表达量较对照分别上调1.146倍、1.379倍、1.212倍;而在根与茎中,其对照表达较叶中高,但低温锻炼下表达上调倍数较叶中低、最高表达量较对照分别上调了1.176倍与1.101倍。另外一种转录因子WRKY在小桐子不同组织中表达差异性与NAC基本一致(图2; 图3),且与文献报道一致(Huang and Duman, 2002; Mare et al., 2004)。该WRKY因子的表达存在显著的组织特异性,其对照表达量的高低顺序为茎、根、叶,但低温锻炼下在茎中的表达差异性较根与叶中要小,在后两者中低温锻炼24h时其表达量都达到了最高值,分别较对照上调了1.284倍与1.052倍。以上结果表明转录因子NAC、WRKY与小桐子抗冷性可能存在密切的关联。
3材料与方法
3.1小桐子材料及处理
供试小桐子种子取自云南省楚雄州元谋县。选取饱满的小桐子种子,用1.5% CuSO4消毒20 min,无菌水漂洗5次,于26℃的恒温培养箱中吸涨24 h (李忠光和龚明, 2010)。将吸涨的种子在无菌水中漂洗3次,播于垫有5层用无菌水湿润滤纸的白磁盘中,于相对湿度(RH) 75%、26/20℃、16/8 h光周期的恒温培养箱中萌发5 d;然后将发芽的种子播于消毒的培养土中,并于同样条件下的恒温培养箱中生长15天至第二片真叶展开,每天用无菌水润湿培养土。将生长15 d的小桐子幼苗置于相对湿度(RH) 75%、12℃、16/8 h光周期的低温培养箱中进行低温锻炼处理,分别取低温锻炼12 h、24 h、48 h与对照(正常培养)的根、茎及第二片真叶,用铝箔纸包好,液氮速冻后于-80℃冰箱中保存用于RNA的提取。
3.2主要试剂
所、TransZol Up、DNase I、TransStart Taq DNA Polymerase、TransScript Two-Step RT-PCR SuperMix、Trans 2K Plus II DNA Marker购自北京全式金生物技术有限公司,其它试剂均为国产分析纯。
3.3引物的设计
使用软件Primer Premier (Version: 5.0)与Vector NTI Advance (Version: 11.5)进行RT-PCR的引物设计,并采用Oligo (Version: 6.0)进行引物验证。设计的引物退火温度控制在49~55℃,扩增片段长度范围为150~500 bp。引物序列见表2,由华大基因公司合成。
3.4试验方法
3.4.1小桐子总RNA的提取及cDNA第一链的合成
称取0.2 g小桐子植物材料,利用TransZol Up试剂提取对照与12℃低温锻炼12 h、24 h、48 h的根、茎及叶片的总RNA,并利用DNase I消化RNA中的基因组DNA,得到纯化的总RNA。分光光度法测定其OD260、OD280、OD230值,计算RNA样品的纯度与浓度。分别取3 μg总RNA,以Anchored Oligo (dT)18为反转录引物,利用TransScript Two-Step RT-PCR SuperMix反转录合成第一链cDNA,-20℃保存。
3.4.2半定量RT-PCR分析
利用小桐子各组织及低温锻炼条件下的反转录cDNA为模板,以18S rRNA为内参,进行各基因的RT-PCR扩增。18S rRNA的扩增条件为:94℃ 5 min→[94℃ 30 s, 55℃ 30 s, 72℃ 1 min]26→72℃ 10 min;其它基因的扩增条件为:94℃ 5min→[94℃ 30 s, T 30 s, 72℃ 1 min]28→72℃ 10 min (退火温度见表2)。反应完毕后取5μl进行1%的琼脂糖凝胶电泳检测。以18S rRNA为比对内参进行各基因表达差异性的分析。利用条带分析软件Quantity ONE计算不同基因在不同组织中于对照和12℃低温锻炼(12 h, 24 h, 48 h)条件下的相对表达量。
3.4.3基因表达模式聚类分析
将上述3.4.2中得到的基因相对表达量数据在Excel软件中规范化成数据矩阵,通过函数计算不同低温处理与对照的比值。将数据导入Cluster 3.0软件,进行以2为底的对数转化,并设置同时以基因与实验处理进行聚类,聚类方法选择K-means,将转化数据导出为聚类分析文件(.cdt文件)。将.cdt文件数据在R软件中进行聚类分析图形绘制,采用的函数为gplots,程序命令为:
> data=read.table("F://R/starch.txt")
> data=as.matrix(data)
> data
> library(gplots)
> heatmap.2(data, col=redgreen(75), scale="none", key=TRUE, symkey=FALSE, density.info= "none" , trace="none", cexRow=0.9, cexCol=0.9)
3.5实验设计和统计分析
试验采取完全随机实验设计(CRD)。数据统计分析使用方差分析技术(ANOVA)。每个处理有20组,三个重复。所有百分比数据进行arcsin比例转换,之后再进行方差分析。
作者贡献
第一作者王海波,主要负责实验工作、数据的处理、论文的撰写及整体修改;龚明与邹竹荣对论文整体结构,以及中英文摘要提供修改意见。
致谢
诚本研究由国家自然科学基金(31260064; 31160169; 31460059; 31460179)和云南省教育厅科研基金重大专项项目(ZD2010004)共同资助。感谢杨玉梅以及实验室的其他同事在中英文资料的收集、前期研究资料提供的帮助。
Ao P.X., Li Z.G., Fan D.M., and Gong M., 2013a, Involvement of antioxidant defense system in chill hardening-induced chilling tolerance in Jatropha Curcas seedlings, Acta Physiol. Plant, 35(1): 153-160
Ao P.X., Li Z.G., and Gong M., 2013b, Involvement of compatible solutes in chill hardening-induced chilling tolerance in Jatropha curcas seedlings, Acta Physiol. Plant, 35(12): 3457-3464
Ferrer J.L., Jez J.M., Bowman M.E., Dixon R.A., and Noel J.P., 1999, Structure of chalcone synthase and the molecular basis of plant polyketide biosynthesis, Nat. Struct. Biol., 6(8): 775-784
Foyer C.H., and Noctor G., 2009, Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implication, Antioxid Redox Signa, 11(4): 861-905
He L., Yu H., Fan Y.H., Sha Y.C., and Yuan L.C., 2010, Plant research progress of Jatropha curcas L., Changjiang Liuyu Ziyuan Yu Huanjing (Resources and Environment in the Yangtza Basin), 19(S1): 120-127 (何璐, 虞泓, 范源洪, 沙毓沧, 袁理春, 2010, 麻疯树(Jatropha curcas L.)植物学研究进展, 长江流域资源与环境, 19 (s1): 120-127)
Huang T., and Duman J.G., 2002, Cloning and characterization of a thermal hysteresis (antifreeze) protein with DNA-binding activity from winter bitter sweet night shade, Solanum dulcamara, Plant Mol. Biol., 48(4): 339-350
Ko J.H., Kim B.G., Hur H.G., Lim Y., and Ahn J.H., 2006, Molecular cloning, expression and characterization of a glycosyltransferase from rice, Plant Cell Rep., 25(7): 141-146
Kodama H., Nishiuchi T., Seo S., Ohashi Y., and Lba K., 2000, Possible involvement of protein phosphorylation in the wound-responsive expression of Arabidopsis plastid omega-3 fatty acid desaturase gene, Plant Sci., 155(2): 153-160
Li Z.G., and Gong M., 2011, Effects of different chemical disinfectant on seed germination and seedling growth of Jatropha curcas L., Zhongzi (Seed), 30(2): 4-7, 12 (李忠光, 龚明, 2010, 不同化学消毒剂对小桐子种子萌发和幼苗生长的影响, 种子, 30(2): 4-7, 12)
Lin J., Zhou X., Tang K.X., and Chen F., 2004, A survey of the studies on the resources of Jatropha curcas L., J. Trop. Subtrop. Bot., 12(3): 285-290
Lorenc-Kukula K., Amarowicz R., Oszmianski J., Doermann P., Starzycki M., Skana J., Zuk M., Kulma A., and Szopa J., 2005, Pleiotropic effect of phenolic compounds content increases in transgenic flax plant, J. Agric. Food Chem., 53(9): 3685-3692
Lumb R.A., and Bulleid N.J., 2002, Is protein disulfide isomerase a redox-dependent molecular chaperone, The EMBO Journal, 21(24): 6763-6770
Mare C., Mazzucotelli E., Crosatti C., Francia E., Stanca A.M., and Cattivelli L., 2004, Hv-WRKY38: A new transcription factor involved in cold- and drought-response in barley, Plant Mol Biol, 55(3): 399-416
Roxas V.P., Lodhi S.A., Garrett D.K., Mahan J.R., and Allen R.D., 2000, Stress tolerance in transgenic tobacco seedling that over express glutathione-S-transferase/glutathione peroxidase, Plant and Cell Physiology, 41(11): 1229-1234
Von Rad U., Huttl R., Lottspeich F., Gierl A., and Frey M., 2001, Two glucosyltransferases are involved in detoxification of benzoxazinoids in maize, Plant J., 28(6): 633-642
Wang H.B., Zou Z.R., Wang S.S., and Gong M., 2014, Deep sequencing-based transcriptome analysis of the oil-bearing plant physic nut (Jatropha curcas L.) under cold treatments, Plant Omics Journal, 7(3): 178-187
Wang H.B., Zou Z.R., Wang S.S., and Gong M., 2013, Global analysis of transcriptome responses and gene expression profiles to cold stress of Jatropha curcas L., PLoS One, 12: e82817
Yu C., Wang H.S., Yang S., Tang X.F., Duan M., and Meng Q.W., 2009, Overexpression of endoplasmic reticulum omega-3 fatty acid desaturase gene improves chilling tolerance in tomato, Plant Physiol. Biochem., 47(11-12): 1102-1112
Zeng J.M., 2006, To developing biomass energy plant—Jatropha curcas L., Yunan Linye (Yunan Forestry), 27(2): 21-22 (曾觉民, 2006, 可大力发展的生物质能源植物——膏桐, 云南林业, 27(2): 21-22)
Zheng X.N., Chen B., Lu G.J., and Han B., 2009, Overexpression of a NAC transcription factor enhances rice drought and salt tolerance, Biochem. Biophys. Res. Commun., 379(4): 985-989