杜鹃花类菌根真菌硝态氮代谢SSH文库构建及分析 
2. 北京林业大学园林学院, 国家花卉工程技术研究中心, 北京, 100083
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2012 年, 第 10 卷, 第 52 篇 doi: 10.5376/mpb.cn.2012.10.0052
收稿日期: 2012年09月03日 接受日期: 2012年10月19日 发表日期: 2012年12月01日
引用格式(中文):
孙秋玲等, 2012, 杜鹃花类菌根真菌硝态氮代谢SSH文库构建及分析, 分子植物育种(online) Vol.10 No.52 pp.1383-1389 (doi: 10.5376/mpb.cn.2012.10.0052)
引用格式(英文):
Sun et al., 2012, Construction and Analysis of SSH cDNA Library of Ericoid Mycorrhizae Fungus Under Nitrate Induction, Fenzi Zhiwu Yuzhong (online) (Molecular Plant Breeding) Vol.10 No.52 pp.1383-1389 (doi: 10.5376/mpb.cn.2012.10.0052)
本研究选取铵态氮下生长及硝态氮诱导的杜鹃花类菌根真菌为材料,利用抑制性差减杂交技术成功构建了硝态氮诱导下杜鹃花类菌根真菌的正向差减cDNA文库。挑取300个阳性克隆进行测序,获得223条单一序列。经Blast分析,其中92条序列为功能已知序列,53条序列为未知功能序列,23条序列能够找到相似性较高基因;55条序列未找到匹配基因。功能已知基因涉及能量代谢、物质合成与代谢、抗性、物质转运、信号转导等代谢活动。其中发现了硝酸盐转运蛋白、硝酸还原酶、亚硝酸还原酶、氨基酸转运蛋白、磷酸转移酶等硝酸盐代谢相关基因,为进一步研究ERM真菌硝态氮代谢提供了分子基础。
氮作为植物的生命元素,不仅是生物有机体的组成成分,还参与酶及叶绿素的合成,植物的生长和产量完全依赖于土壤中的氮含量,尤其是硝态氮、铵态氮等有效氮含量(Gobert and Plassard, 2008)。增强植物对于氮素的利用能力和效率,对于植物生长和整个生态系统的稳定性有重要的意义。大多数杜鹃花科植物自然生境土壤为酸性土,通常认为在pH低于5.5的土壤中硝化作用几乎是可以忽略不计的(Paul and Clark, 1989),酸性土壤中硝态氮不可利用(Kosola and Workmaster, 2007)。而1919-1971年间所发表的有关杜鹃花科重要植物蓝莓(Vaccinium macrocarpon)的研究论文均认为:当施以硝态氮时,蓝莓的生长量比无氮源条件下大,甚至有试验发现蓝莓在以硝态氮为氮源的基质中生长量大于以铵盐为氮源的生长量(Kender and Childers, 1959)。由于蓝莓根系自然状态下均能形成杜鹃花类菌根(ericoid mycorrhizas, ERM),Read (1996)推测菌根可能增强了寄主植物利用硝态氮的能力(Read, 1996)。此后,经过ERM真菌纯培养以及接种蓝莓(Kosola and Workmaster, 2007)、云锦杜鹃(Rhododendron fortunei L.) (尹丽娟等, 2010)试验进一步证明ERM真菌具有吸收利用硝态氮的能力。但研究者未曾就ERM真菌吸收硝态氮的机制进行深入研究。
抑制性差减杂交(suppression subtractive hybridization,SSH)自1996年由Diatchnoko等发明以来,由于其具备特异性高、假阳性率低,对低丰度基因有较好的分离效果等优点,被广泛用于分离各类生物体的差异表达基因,这些基因主要涉及生命活动包括胚胎发育成花等机体发育(黄鑫, 2008; 侯雪丽, 2009)、形态建成(王永胜等, 2001; 张雷等, 2002, 自然科学进展, 12(3): 261-265)、生物与非生物胁迫(申贵, 2005)等。
本研究以ERM真菌中最为普遍的一类真菌为试验材料,利用抑制性差减杂交(suppression subtractive hybridization, SSH)首次构建了ERM真菌硝态氮诱导下的cDNA文库,以期从中得到ERM真菌硝态氮代谢相关的基因序列,为揭示ERM硝态氮代谢机理提供参考资料。
1结果分析
1.1总RNA质量检测及mRNA的分离纯化
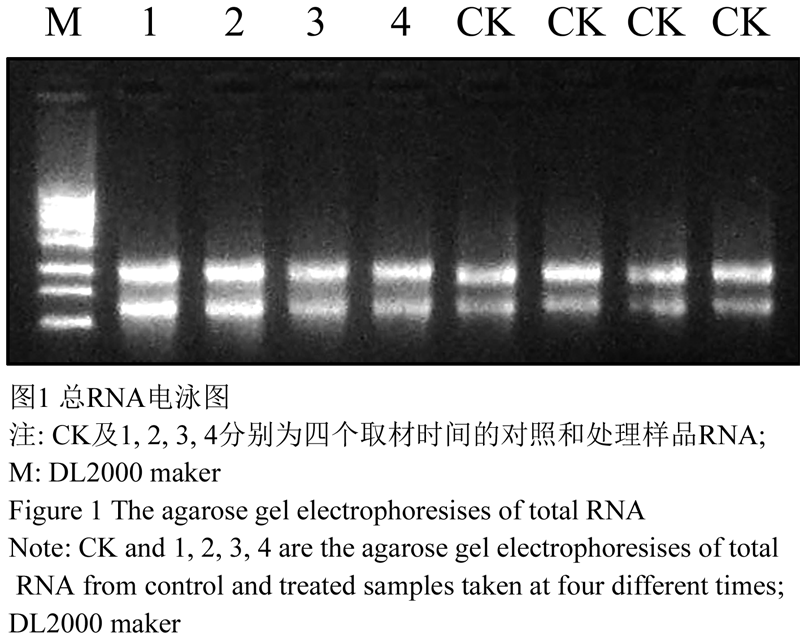
提取的总RNA 28S和18S带型清晰,与EB结合量比值接近2:1,且两条带之间未出现弥散现象,几乎没有DNA污染(图1)。对照和处理组总RNA OD260/OD280分别为1.900和1.860,介于1.7~2.0之间(表1),说明所提RNA样品受多糖、蛋白质等物质污染程度较小,质量较好,纯度高。分离的mRNA在琼脂糖凝胶上呈均匀的弥散状(图2),说明采用Promega公司的PolyA Tract SystemsⅢ (9Z5300)纯化出的mRNA具有较好的完整性,未被降解,可用于后续文库构建。
|
|
|
|
.jpg)
|
|
.png)
1.2 SSH文库构建效果检测
1.2.1 RsaⅠ酶切效率分析
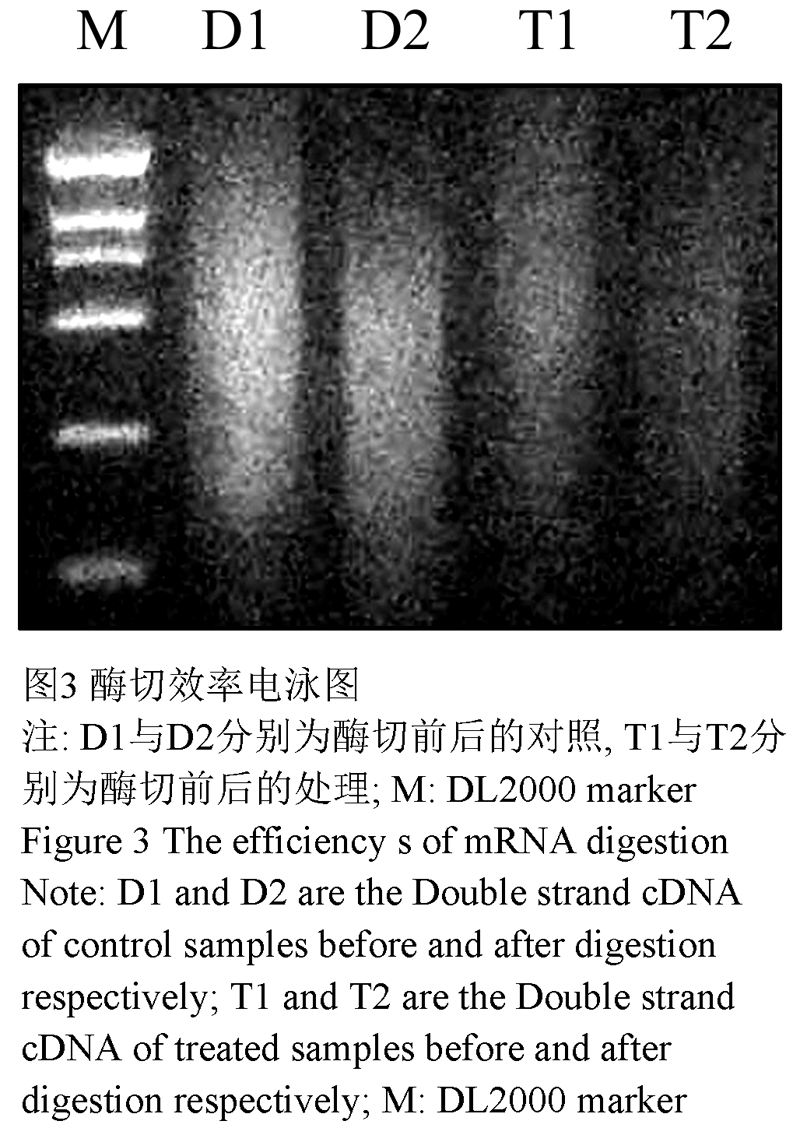
取处理组和对照组mRNA各2 µg,反转录成双链cDNA后,经RsaⅠ酶切,并用2%琼脂糖/EB胶电泳检测酶切效率(图3),处理组和对照组的完整cDNA从100 bp到点样孔成弥散状分布,主要集中在150~2 000 bp以上,酶切后cDNA较酶切前分布范围明显下移,亮度主要集中在200~700 bp之间,说明酶切效果较好,可用于后续接头的连接和差减杂交。
|
|
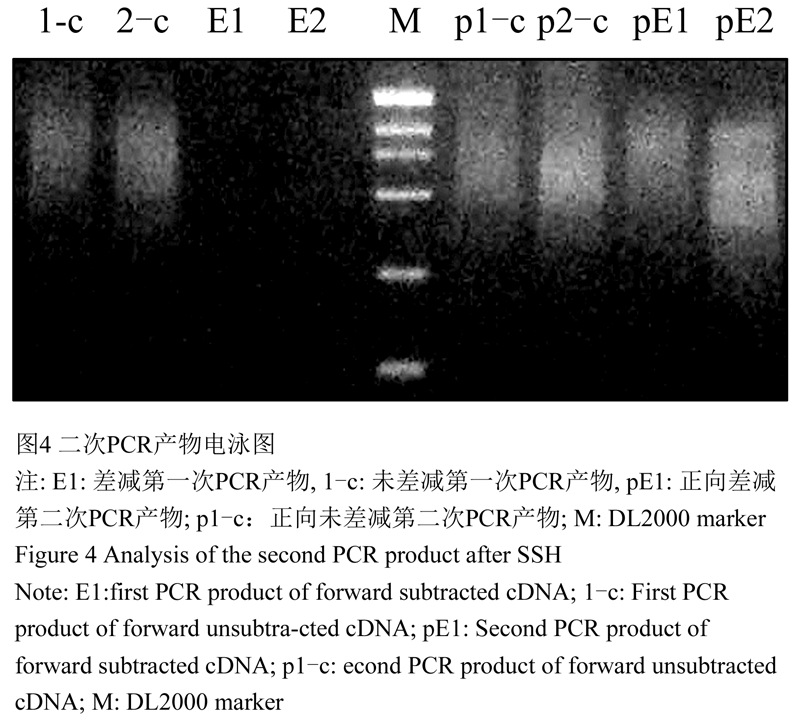
1.2.2 抑制性差减杂交(SSH)后二次PCR产物分析
差减片段经两轮PCR扩增后,二次PCR产物电泳结果(图4)显示,差异片段得到了有效富集,扩增cDNA片段成弥散状分布,未见明显特异性条带,说明抑制性差减杂交和PCR扩增效果较好。
|
|
1.2.3 文库插入片段的大小检测
随机挑选46个克隆进行PCR扩增,检测所构建文库中插入片段的大小,结果如图5所示:插入片段最小为200 bp,最大为1 000 bp,大部分集中在400 bp左右。表明连接、转化效果较好,片段大小符合文库构建质量要求。
|
|
1.2.4 cDNA文库中ESTs序列及功能分析
300个阳性克隆测序完成后,共有270个克隆测序成功,去除低质量序列、小于100 bp序列以及冗余序列后,得到223条单一序列。序列最短为181 bp,最长1 007 bp,平均509 bp,绝大多数集中在300~600 bp之间,与PCR检测结果相一致。
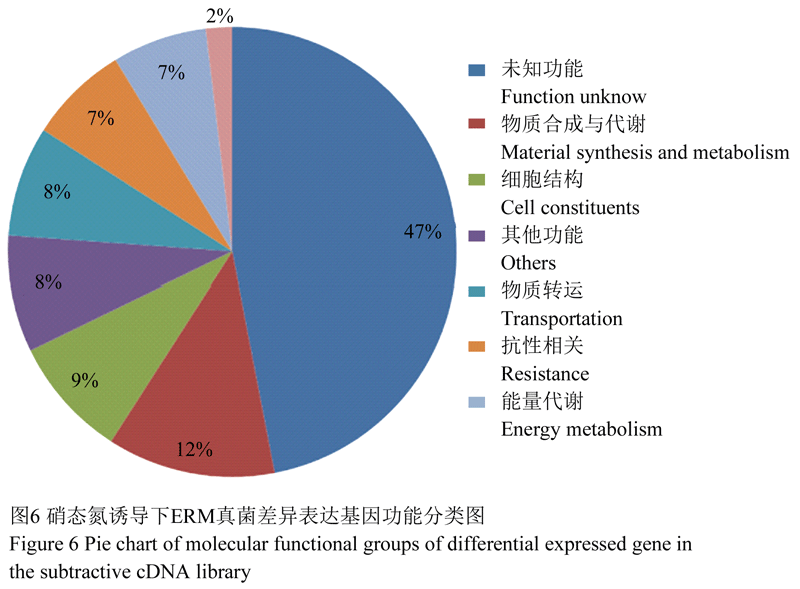
经Blast分析,发现223条序列中92条为功能已知序列,53条为未知功能序列,分别占全部基因的41.26%和23.77%。23条序列能够找到相似性较高基因;55条序列未找到匹配基因,占到全部基因的24.66%。为进一步了解文库中ESTs所涉及的生命活动,结合本试验的研究目的,将所得全部ESTs具体分为八类,包括能量代谢、物质合成与代谢、抗性、物质转运、细胞结构、信号转导、其他功能、未知功能。按照基因在所得文库中所占的比例,绘制饼状图(图6)。未知功能基因所占比例最大,占到了全部基因的47%;物质合成代谢相关ESTSS在功能已知基因序列中所占比列最大,为12%,其中包含与硝态氮代谢相关基因有硝酸还原酶、亚硝酸还原酶基因、氨基酸合成酶等,除此还涉及到基因的转录和蛋白质的合成等生命活动;细胞结构基因所占比例仅次于物质代谢基因,其中包含60s核糖体蛋白、组蛋白、核仁蛋白、染色体、GPI锚定蛋白等;物质转运相关基因所占比例8%,其中包含多种物质的转运蛋白,如:硝态氮转运蛋白(NRT)、K离子通道、氨基酸转移酶、磷酸转运酶以及MFS单糖转运蛋白等;能量代谢相关基因占到了全部基因的7%,其中包含丙酮酸脱羧酶、糖原合成酶、异柠檬酸裂合酶、GTP环水解酶、糖基转移酶、ATP伴侣蛋白等;其它还有抗性相关基因包含一些热激蛋白、细胞色素C氧化酶、以及泛素代谢相关基因等。表2显示的是部分所得片段的Blast结果。
|
|
|
|
.jpg)
2讨论
抑制性差减杂交理论上只适用于两个样本间的差异比较分析,且要求样本间差异不可过大或过小。本试验中,ERM真菌菌株19在初始浓度为3.8 mmol/L的MMN培养液中已生长15 d,其生长所产生的次级代谢物势必导致培养液成分的变化,且氮源终浓度不确定,为了排除硝态氮诱导培养液与对照培养液成分差距过大,保证两种培养液的差别在可控制范围内,本试验在菌株培养结束时,先将菌丝转入新鲜铵态氮培养液重新培养24 h,之后收集一部分菌丝作为对照,另一部分转入硝态氮培养液中培养作为试验方,最终成功获得了硝态氮代谢相关基因,说明该处理方法比较合理。
从所建文库中,我们共获得3条硝态氮转运蛋白基因、一条硝酸还原酶基因和一条亚硝酸还原酶基因,同时还得到3条氨基酸转运蛋白基因以及其它与物质合成代谢相关的基因序列。其中硝酸盐转运蛋白负责将外界硝酸盐转运至生物体内,这是生物体硝酸盐利用的第一步。植物体内的硝酸盐转运蛋白通常以多个成员组成的多基因家族的形式存在,依据生理特性不同,被划分为高亲和性转运系统(high-affinity transport systems, HATS)和低亲和性转运系统(low-affinity transport systems, LATS),当外界硝酸盐浓度较高时(>1 mmol/L),低亲和性转运系统起作用,当浓度较低时(<250 μmol/L),高亲和性转运系统起作用。本试验氮浓度设为100 μmol/L,硝酸盐转运蛋白基因共出现6次,说明此低浓度氮素条件下该基因的表达量很高,推测次ERM真菌的NRT属于高亲和转运系统家族成员之一,这仍需要生理和分子试验证据证明。由于硝酸盐中的氮为高度氧化态氮,在参与合成体内多数还原态氮化合物之前,必须先被还原为还原态的氨,而硝酸还原酶和亚硝酸还原酶是该过程中的关键酶和限速酶,硝酸还原酶负责将硝酸盐还原为亚硝酸,亚硝酸经亚硝酸还原酶再还原为铵盐。菌根真菌中还原为铵盐之前的代谢过程很少被研究,但最近几年也引起了研究者的注意,主要围绕的是外生菌根真菌中上述基因的表达进行,证明了该类基因与植物基因不同,它不仅受硝酸盐的诱导,无氮条件下其依然上调表达(Jargeat et al., 2003; Guescini et al., 2007),ERM真菌中该类基因的表达模式与此是否相同还需进行进一步的试验验证。铵盐后续代谢在AM菌根中的研究比较深入,外生菌根真菌也有涉及,已被证实AM菌根真菌中铵盐的后续代谢是一系列酶促反应的过程,其中涉及到多种氨基酸的合成和转运(Govindarajulu et al., 2005; Montanini et al., 2006),本研究中氨基酸转运蛋白共出现4次,共获得3条不同ESTs序列;精氨酸是AM真菌氮素代谢的关键中间产物,是氮素从根外菌丝进入根内菌丝的主要氮素形态,而精氨酸很可能是与多聚磷酸盐一起转运完成氮素从根外菌丝转至根内菌丝的过程(Guescini et al., 2007; Cruz et al., 2007),本研究中也获得两条磷酸转移酶的EST序列,这些都说明ERM真菌可能存在与AM菌根真菌同样的精氨酸转运模式。研究中获得了很多未知基因,已知基因也大多数没有明确的功能注释,ERM真菌利用硝态氮的代谢途径还远未揭示,有待进一步研究。今后将针对筛选出的重要差异表达基因采用实时定量PCR和Northern法进一步进行验证, 并进行全长基因的克隆及其表达和功能分析,逐步揭示ERM真菌硝态氮利用的机制,最终服务于植物菌根技术的应用。
3材料与方法
3.1材料
试验选取在硝态氮培养基上生长良好的ERM真菌 “菌株19”作为试验材料。该菌株为子囊菌门(Ascomycota)、锤舌菌纲(Leotiomycetes)、粘毛菌科(Myxotrichaceae)、树粉孢属(Oidiodendron)真菌菌株,分离自浙江华顶山采集的云锦杜鹃根系。
试验所用生化试剂均为国产分析纯,购自国药集团;常用分子生物学试剂均购于鼎国昌盛生物技术有限公司;PolyA Tract SystemsⅢ(Z5300)购于Promega公司,PCR-selectTM cDNA subtraction kit以及AdvantageTM cDNA Polymerase Mix购于Clontech公司,pMD-19T载体购于TAKARA公司。
试验菌丝培养所用基本培养液为MMN (Melin-Norkrans Medium)培养液,去除其中的麦芽提取物,所用氮源为(NH4)2HPO4,终浓度3.8 mmol/L,转速150 r/min摇床内连续暗培养15 d,温度为(25±1)℃。
3.2方法
3.2.1 试验材料处理
菌株培养15 d后,过滤收集菌丝,重悬于新鲜MMN培养液中振荡培养24 h,再次过滤,一半重悬于新鲜MMN培养液中振荡培养,作为对照组;一半重悬于以100 µmol/L Ca(NO3)2为唯一氮源的MMN培养液中,进行硝态氮诱导,作为处理组,分别振荡培养4 h、8 h、24 h、36 h取材,液氮速存。
3.2.2 总RNA的提取及mRNA的纯化
菌丝总RNA利用鼎国公司的PlantRNAzol试剂盒提取,步骤同试剂盒说明书。用1%琼脂糖凝胶电泳,检测RNA提取质量,并用微量紫外分光光度计测定其纯度和浓度;采用Promega公司的PolyA Tract SystemsⅢ(Z5300)分离mRNA,并利用琼脂糖凝胶电泳检测其完整性。
3.2.3 抑制性差减杂交
采用Clontech公司的PCR-selectTM cDNA Subtraction Kit试剂盒,并按其说明书进行。
3.2.4 差减cDNN文库的构建及序列分析
第二轮PCR扩增结束后,将产物与载体pMD-19T连接,再转化大肠杆菌感受态细胞DH5a,完成cDNA文库构建。随机选择300个阳性克隆进行测序,去除测序序列中的载体序列和接头序列后,即得EST序列,于NCBI上进行Blast同源比对及功能注释后,将所得ESTs进行基因功能分类。
作者贡献
张春英、孙秋玲是本研究的实验设计和实验研究的执行人,张春英是项目的构思者和负责人,指导实验设计,数据分析,论文写作与修改;戴思兰指导论文的写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由国家自然科学基金项目(30972409)资助。作者感谢上海市园林科学研究所尹丽娟、魏翔莺、黄芳以及师姐陈真在本实验过程中的帮助。感谢两位匿名的同行评审人的评审建议和修改建议。
参考文献
Cruz C., Egsgaard H., Trujillo C., Ambus P., Requena N., Martins-Loução M.A., and Jakobsen I., 2007. Enzymatic evidence for the key role of arginine in nitrogen translocation by arbuscular mycorrhizal fungi. Plant Physiol, 144(2): 782-792.
http://dx.doi.org/10.1104/pp.106.090522 PMid:17142485 PMCid:1914155
Gobert A.,and Plassard C., 2008 The beneficial effect of mycorrhizae on N utilization by the host-plant: myth or reality? In: Mycorrhiza, springer, GER, pp.209-240.
http://dx.doi.org/10.1007/978-3-540-78826-3_11
Guescini M., Zeppa S., and Pierleonl R., 2007 The expression profile of the Tuber borchii nitrite reductase suggest its positive contribution to host plant nitrogen nutrition. Curr. Genet, , 51(1): 31−41.
http://dx.doi.org/10.1007/s00294-006-0105-y PMid:17082947
Hou X.L., 2009, Studies on the differentially expressed genes in accessory sex gland and function of eriocheir sinensis, Thesis for M.S., East China Normal University, Supervisor: Wang Q., PP.1-74 (侯雪丽, 2009, 中华绒鳌蟹副性腺差减cDNA文库的构建及内容物功能的研究, 硕士学业论文, 华东师范大学, 导师: 王群, pp.1-74)
Huang X., 2008, Cloning and analysis of genes associated with the release of dormant floral buds in tree peony (Paeonia suffruticosa Andr.), Thesis for Ph.D., Beijing Forestry University, Supervisor: Zheng G.S., Dai S.L., pp.1-104 (黄鑫, 2008, 牡丹花芽内休眠解除相关基因的分离与功能分析, 博士学位论文, 北京林业大学, 导师: 郑国生, 戴思兰, pp.1-104)
Jargeat P., Rekangalt D., Verner M.C., Gay G., Debaud J.C., Marmeisse R., and Fraissinet-Tachet L., 2003, Characterisation and expression analysis of a nitrate transporter and nitrite reductase genes, two members of a gene cluster for nitrate assimilation from the symbiotic basidiomycete Hebeloma cylindrosporum. Curr. Genet, 43(3): 199-205.
Kender W.J., and Childers NF. 1959. Growth of cranberry plants (Vaccinium macrocarpon) with various sources of nitrogen. Proc. Natl. Acad. Sci. U S A.,. 74: 407–413.
Kosola K.R., and Workmaster B.A.A.. 2007. Mycorrhizal colonization of cranberry effects of cultivar, soil type, and leaf litter composition. J. Am. Soc. Hortic. Sci. 132(1): 134-141
Govindarajulu M., Pfeffer P.E., Jin H., Abubaker J., Douds D.D., Allen J.W., Bücking H., Lammers P.J., and Shachar-Hill Y., 2005. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature, 435(7043): 819-823.
http://dx.doi.org/10.1038/nature03610 PMid:15944705
Montanini B., Viscomi A.R., Bolchi A., Martin Y., Siverio J.M, Balestrini R., Bonfante P., and Ottonello S., 2006, Functional properties and differential mode of regulation of the nitrate transporter from a plant symbiotic ascomycete. Biochem. J, 394(1): 125-134.
http://dx.doi.org/10.1042/BJ20051199 PMid:16201972 PMCid:1386010
Paul E.A., and Clark F.E., 1989, Soil microbiology and biochemistry, Academic Press, San Diego, CA, USA, pp.273
Read D.J., 1996, The structure and function of the ericoid mycorrhizal root, Ann. Bot, 77(4): 365-374.
http://dx.doi.org/10.1006/anbo.1996.0044
Shen G., 2005, Differentially expressed genes during early infection of Phytophthora Sojae with soybean plant and functional analysis of some genes, Thesis for Ph.D., Nanjing Agricultural University, Supervisor: Zheng X.B., PP.1-158 (申贵, 2005, 大豆疫霉侵染大豆早期差异表达基因的筛选与功能分析, 博士学位论文, 南京农业大学, 导师: 郑小波, pp.1-158)
Wang Y.S., Wang J., Li F.Q., Liu X.B., Liu L.S., 2001, Identification of cDNA fragments differentially expressed between a rice mutant dwarf69 and wild type TGA-2 suppressive subtraction hybridization, GaoJishu Tongxun (High Technology Letters), 5: 20-24 (王永胜, 王景, 李发强, 刘筱斌, 刘良式, 2001, SSH法获取水稻矮化突变体相关的cDNA片段, 高技术通讯,11(5): 20-24)
Yin L.J., Zhang C.Y., Yang B., 2010, Characteristics of nitrogen absorbed by Ericoid Mycorrhizal fungi and impact on growth of Rhododendron fortunei, Zhongguo Nongye Kexue (Scientia Agricultura Sinica), 43(2):868-872 (尹丽娟, 张春英, 杨兵, 2010, 云锦杜鹃菌根真菌吸收氮源特性及其接种效应, 中国农业科学, 43(4): 868−872)