贵州辣椒疫霉菌交配型测定及遗传多样性分析 
2. 贵州省植物保护研究所, 贵阳, 550006
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2013 年, 第 11 卷, 第 17 篇 doi: 10.5376/mpb.cn.2013.11.0017
收稿日期: 2013年03月13日 接受日期: 2013年04月29日 发表日期: 2013年06月14日
引用格式(中文):
陈龙等, 2013, 贵州辣椒疫霉菌交配型测定及遗传多样性分析, 分子植物育种(online), 11(17): 1114-1122 (doi: 10.5376/mpb.cn.2013.11.0017)
引用格式(英文):
Chen et al., 2013, Mating Type Determination and Genetic Diversity of Phytophthora capsici Isolates in Guizhou, Fenzi Zhiwu Yuzhong (online) (Molecular Plant Breeding), 11(17): 1114-1122 (doi: 10.5376/mpb.cn.2013.11.0017)
采用单孢菌落直接配对法和RAPD分子标记对源自贵州省贵阳、毕节、遵义、黔南、安顺和六盘水等6个地区10余个县乡的的72个辣椒疫霉单孢菌株分别进行交配型测定和遗传多样性分析。结果表明,72个菌株分属A1、A2和A1,A2三种交配型,发生频率分别为8.3%、88.9%、2.8%,以A2交配型占绝对优势,在各产区均有分布。RAPD遗传多样性分析显示,筛选的31个引物共标记出329条DNA指纹条带,其中317条具有多态性,多态性检测率为96.35%,各地区菌株Shannon's多样性指数在0.1142~0.4244之间,平均为0.2224;其顺序为:贵阳(0.4244)>遵义(0.2995)>毕节(0.1926)>黔南(0.1822)>安顺(0.1206)>六盘水(0.1142),表明贵州辣椒疫霉菌具有丰富的遗传多样性。以遗传相似系数0.78为阈值,UPGMA聚类分析法将供试菌株划分为7大类群,聚类组的划分与菌株地理分布、寄主品种显示一定的相关性。
辣椒疫病是由辣椒疫霉菌(Phytophthora capsici Leonian)侵染引起的一种土传真菌病害(Hausbeck et al., 2004)。除侵染辣椒外,该病原菌还可引起茄子、番茄、黄瓜等多种重要蔬菜作物的疫病(王志田, 1988, 新疆农业科学, (2): 27-28; 文景芝等, 1998)。在中国,该病发生最为严重的地区为江苏、安徽、浙江和上海等长江中下游省份(兰海等, 2008, 湖北植保, (1): 16-17)。自杨学辉(2004)首次报道贵州辣椒疫病病害以来,该病害在贵州的辣椒产区也有逐年加重的趋势。
P. capsici存在性分化现象,属于异宗配合的卵菌,不同交配型的菌株相互诱导能产生具有很强抗逆能力的卵孢子,使物种得以延续;而且两个可亲和菌系配合时可能发生基因重组,使该病原菌获得更强的生存能力、致病力和更广泛的寄主范围(何汉兴等, 1983; Lamour and Hausbeck, 2001)。研究表明,疫霉菌包含5种交配型(郑小波, 1997, 中国农业出版社, pp.30-33),以A1和A2为主,其分布频率常处于动态变化之中并有一定的地域分布特征(Goodwin and Drenth, 1997; Islam et al., 2005; Silvar et al., 2006; 郑小波和陆家云, 1990)。刘永刚等(2008)、杨明英等(2009)对中国甘肃、云南等省辣椒疫霉菌株交配型组成和分布状况的研究表明,各地区辣椒疫霉交配型的构成不尽相同,其分布有明显的地理差异。
随着遗传学及分子生物学技术的发展,RAPD等分子标记技术已被广泛用于物种多样性及种群间亲缘关系的研究。Silvar等(2006)利用RAPD对西班牙、巴西、伊朗、印度等国家的辣椒疫霉菌株进行遗传多样性分析,发现西班牙菌株具有较低的遗传变异,来自不同国家的菌株间遗传差异非常明显,地理位置越接近的国家之间菌株遗传相似性越高。李萍等(2011)对来自我国安徽、江苏及四川等县市的辣椒疫霉菌进行了RAPD遗传多样性分析,结果表明辣椒疫霉菌株遗传多样性较丰富,亲缘关系的远近与地理来源有一定相关性。
贵州是中国辣椒主产区,辣椒疫病的危害已成为产业发展的障碍之一,为了做好该病的科学防控,有必要明确本地辣椒疫霉菌的种群组成及其变异特点,为此,本文分别采用单胞菌落直接配对法和RAPD分子标记技术对贵州省不同地区的辣椒疫霉菌株进行了交配型测定和遗传多样性分析,旨在探明贵州辣椒疫霉菌交配型的构成与分布、群体亲缘关系及遗传变异特性,为辣椒疫病综合防治策略的制定提供依据。
1结果与分析
1.1贵州辣椒疫霉交配型组成与分布
供试的72个辣椒疫霉菌由A1、A2和A1,A2三种交配型组成,其中64株为A2交配型,各采集区均有分布,发生频率为88.9%;6株为A1交配型,分布于贵阳、安顺和都匀地区,发生频率为8.3%;2株为A1,A2交配型,分别来自贵阳和都匀,发生频率为2.8%;在所测菌株中未发现A0和A1A2交配型菌株(表1)。这表明贵州省辣椒疫霉菌的交配型有一定多样性,但以A2型占绝对优势,为贵州辣椒产区的优势病原菌群体。
 表1 贵州省辣椒疫霉菌的交配型分布及发生频率 Table 1 Distribution and occurrence frequency of mating types of P. capsici in Guizhou Province |
从辣椒疫霉在各个地区的分布来看,A2交配型菌株仍占绝对优势。其中,遵义、毕节和六盘水地区仅有A 2交配型,发生频率100%;安顺地区发现两种A1、A2两种交配型,频率分别为5.9%、94.1%;贵阳和黔南地区有A1、A2和A1,A2三种交配型菌株分布,出现频率分别为8.3%、88.9%和2.8%。
1.2辣椒疫霉菌的遗传多样性
利用31个RAPD引物对72株辣椒疫霉菌进行扩增,不同引物对供试菌株扩增出的DNA谱带不尽相同,变化幅度为6~17条,平均10.6条;总共扩增出329条谱带,其中317条为多态性标记,多态位点百分率(PPB)为96.35%。供试菌株既具有相同的主扩增带,又有多态性条带,这在一定程度上说明贵州省辣椒疫霉菌既具有种内的同源性,也有其遗传背景的复杂性。
不同地理种群,Nei's基因多样性指数大小范围为0.071 7~0.280 0,平均为0.121 0。Shannon's多样性指数大小范围为0.114 2~0.424 4,平均为0.222 4。六个地理种群的Nei's指数和Shannon's指数的计算结果趋势一致,都以贵阳、遵义较高。Shannon's多样性指数各组大小顺序为:贵阳(0.424 4)>遵义(0.299 5)>毕节(0.192 6)>黔南(0.182 2)>安顺(0.120 6)>六盘水(0.114 2)(表2)。
 表2 不同地理来源辣椒疫霉种群的遗传参数 Table 2 The genetic parameters of Phytophthora capsici populations from different geographical regions |
从不同地理来源上分析辣椒疫霉菌种群间的遗传相似性(表3),安顺和黔南的菌株间遗传相似性最高为0.955 7,其次为六盘水与遵义的菌株间遗传相似性为0.9302、遵义与贵阳的菌株间遗传相似性为0.923 0。遗传相似性较低的为毕节与六盘水、毕节与安顺的菌株,分别为0.747 7、0.752 9。毕节与其他地区的菌株遗传相似性也较低,均低于0.810 0。菌株间的遗传相似性能反应它们之间的亲缘关系,即相似性越高,菌株间的亲缘关系越近。由图1看出,相似性较高的安顺与黔南、六盘水与遵义首先分别聚在一起,毕节与其他地区遗传距离最远,遗传差异最大。
 表3 辣椒疫霉不同地理种群的遗传相似性和遗传距离 Table 3 Genetic identity and genetic distance from different geographical groups of P. capsici |
 图1 不同地区菌株的系统树图 Figure 1 Dendrogram of P. capsici from different regions |
1.3供试菌株RAPD聚类分析
从图2可知,以遗传相似系数0.78为阈值,供试的72个辣椒疫霉菌被划分为7个遗传聚类组。对受试菌株群体聚类组划分进一步分析发现:(1)受试辣椒疫霉菌聚类组的划分与菌株地理来源有一定的关系,但不明显;部分地区的菌株间存在严重的交叉聚集现象。第Ⅰ组的12个菌株中5个来自六盘水,5个来自遵义,2个来自贵阳;第Ⅱ组为优势种群,共计41个菌株,其中4个来自贵阳,17个来自安顺,19个来自黔南,1个来自遵义;第Ⅲ组只有一个菌株,来自贵阳;第Ⅳ组的12个菌株中1个来自六盘水,6个来自遵义,4个来自贵阳,1个来自黔南;第Ⅴ、Ⅵ组分别由2个菌株组成,均来自毕节;第Ⅶ组由来自贵阳的2个菌株组成。(2)受试贵阳地区的13个菌株分别被划分于5个聚类组中,说明贵阳地区的辣椒疫霉菌株变异性较大,遗传分化明显,而且菌株68-2,3-1-1和3-1-2还被分别列为一个独立的聚类组。(3)安顺全部菌株和黔南95%菌株都位于同一聚类组中,可见这些地区的受试菌株分化程度较小,有较近的亲缘关系。(4)毕节地区受试菌株与其他地区菌株的遗传相似系数相对较低,遗传背景较远。该结果与popgene软件统计结果一致。
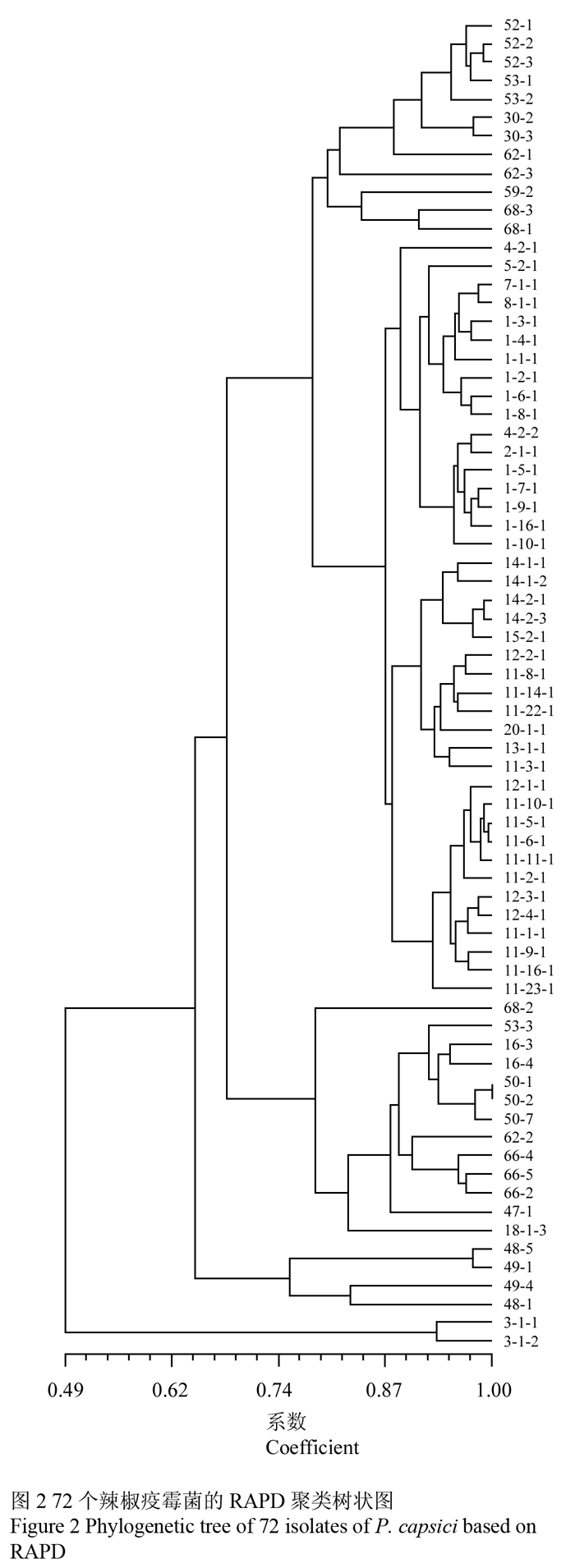 图2 72个辣椒疫霉菌的RAPD聚类树状图 Figure 2 Phylogenetic tree of 72 isolates of P. capsici based on RAPD |
此外,菌株群体聚类组的划分与寄主品种也显示出一定关系(表4),聚类组Ⅲ的菌株来自于辣甜椒;聚类组Ⅴ和Ⅵ的菌株来自于大方皱椒;聚类组Ⅶ的菌株来自于山辣椒。
 表4 贵州辣椒疫霉菌RAPD-PCR扩增结果的UPGMA聚类结果统计表 Table 4 Statistics of 72 P. capsici isolates from Guizhou by UPGMA analysis on RAPD-PCR |
2讨论
本研究的测定结果表明,贵州省辣椒疫霉主要有A1、A2T和A1,A2三种交配型,其分布呈极不均等态势,以A2型占据绝对优势。这与国内其他省份辣椒疫霉的交配型分布有所不同,例如刘永刚等(2008)研究发现甘肃省是以A1型为主,但A0型也占有相当比例;云南省则以自育的A1A2型为主(杨明英等, 2009);安徽省以A1和A2型为主(戚仁德等, 2012);新疆辣椒疫霉有同宗配合现象(贾菊生, 1992)。多种交配型的同时存在意味着辣椒疫霉菌株具有进行有性生殖发生遗传重组的可能,增加了生产上辣椒品种抗性丧失的风险,为病害的发生和流行提供初侵染源、加大了病害的防治难道。此外,笔者还发现,供试72株菌株中,2012年之前(2004年, 2007年, 2009年)采集保存的28株菌株全部为A2交配型,2012年采集分离的44株菌株交配型则有A1、A2和A1,A2三种,产生这种现象的原因可能与近年来贵州辣椒种植面积不断扩大、品种频繁引进有关。辣椒疫病虽是土传病害,但其带菌种子或病残体却可随种子调运进行远距离传播,同时,国外已有研究报道(Noon and Hickman, 1974)有关P. capsici的A2交配型菌株在含地茂散的培养基上可自孕产生卵孢子,转变为A1A2型菌株,这种自孕能力在不含地茂散的培养基上可长久保持。戚仁德等(2012)研究安徽省P. capsici交配型在无性后代的遗传,发现用甲霜灵处理不会诱导P. capsici交配型的变异。至于贵州P. capsici交配型变异及多样化产生原因还需通过诸如种子带菌检测、杀菌剂诱导及定向选择等一系列实验来证实。
从RAPD-PCR分析结果看,贵州省辣椒疫霉菌群体存在丰富的遗传多样性,遗传变异最大、分化最明显的是贵阳地区,亲缘关系最近的是黔南和安顺,毕节跟其他地区遗传背景较远。聚类分析结果表明,聚类组的划分与菌株地理来源及寄主品种显示一定的相关性。这可能是由于:第一,地理位置较近的区域,对于辣椒疫霉菌来说其生存环境较为相近,进化方向差异性小;第二,植物病原菌的致病性变异,是寄主长期压力选择的结果,例如来自安顺的17个菌株全部分离于安椒和陆丰43两个品种,这两个品种已在安顺天龙镇的几个蔬菜基地中种植多年,而分离自毕节的菌株则是来源于当地多年种植的地方品种,结果另文发表。因此,应加强新抗源的引进和广谱抗病品种的选育,进行栽培品种的合理布局和轮换种植,避免因品种单一化种植而引起病害大流行。
3材料与方法
3.1供试材料
供试72个单孢菌株,分别从贵州省遵义、贵阳、安顺、黔南、毕节和六盘水等地区(州, 市)所属10余个县乡的辣椒病株上分离,详见表5。标准菌株A1和A2由中国农业大学刘西莉教授馈赠。
 表5 供试辣椒疫霉菌株来源 Table 5 Sources of the tested isolates of Phytophthora capsici |
3.2交配型测定
将标准交配型菌株及供试菌株分别在CA培养基上培养一周,沿新鲜菌落边缘用灭菌打孔器分别取直径相同的菌丝块,采用单孢菌落直接配对法培养观察性器官的产生情况,方法详见(郑小波, 1997, 中国农业出版社, pp.88-89)。
3.3基因组DNA的提取与RAPD分析
将经过活化的单孢菌株接种于灭菌的CA (200 g胡萝卜榨汁, 蒸馏水1 000 mL)液体培养基、摇床震荡(100 r/min)培养7~10 d,用双层灭菌纱布收集菌丝并置于65℃恒温箱干燥,取出后直接用于DNA提取或-20℃保存备用。辣椒疫霉菌基因组DNA的制备采用生工生物工程有限公司生产的Ezup株式真菌基因组DNA抽提试剂盒提取DNA,用1%琼脂糖胶检测浓度。从上海生工生物工程有限公司所产的100个十核苷酸随机引物中筛选出条带扩增多且清晰的31个随机引物对供试菌株进行PCR扩增。RAPD-PCR反应参考Williams等(1990)的方法,反应体系总体积为25 µL,包含12.5 µL 2×Tap MasterMix、0.4 µmol/L引物和15~20 ng DNA模板。PCR反应在PTC -200仪上进行,热循环反应参数为:94℃预变性2 min,94℃变性1 min,36℃退火30 S、72℃延伸2 min,40个循环,最后72℃延伸10 min,产物4℃保存。凝胶扩增产物在含有GoldviewⅠ的1%琼脂糖凝胶中电泳2.5h,经紫外透射仪观察照相,所有引物均对受试菌株进行3次重复扩增。
3.4数据处理
3.4.1交配型确定
待测菌株的交配型按照如下标准确定:仅与A1交配型配对完成有性生殖的为A2交配型;仅与A2交配型配对完成有性生殖的为A1交配型;与A1、A2交配型配对均能完成有性生殖,但单株培养不能完成有性生殖的为A1,A2交配型;与A1、A2交配型配对均不能完成有性生殖的为A0交配型;单株培养即可完成有性生殖的为A1A2交配型,方法参考(郑小波, 1997, 中国农业出版社, pp.31)。
3.4.2多态性分析
观察每个位点扩增条带的有无,有带记为“1”,无带记为“0”,形成“0,1”数据构成的原始矩阵。利用NTSYS-PC Version 2.11软件计算遗传相似系数,采用非加权组平均法(UPGMA)进行SAHN聚类分析,生成供试菌株间的遗传聚类图;利用Popgene32软件计算群体内及群体间Nei's和Shannon's多样性指数。
作者贡献
陈龙是本文实验研究的主要执行人,并主要完成数据分析及论文写作;袁洁参与实验设计与实验指导;何海永、陈小均、吴石平参与实验材料选择、实验设计及结果分析;杨学辉、王云月是项目的构思者及负责人,指导实验设计,数据分析,论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由贵州省科技攻关项目(黔科合NY字(2009)3002)资助。感谢贵州省植物保护研究所王利爽和谭清群两位女士在实验过程中给予的帮助,同时感谢中国农业大学刘西莉教授馈赠标准型实验菌株。
参考文献
Goodwin S.B., and Drenth A., 1997, Origin of the A2 mating type of phytophthora infestans outside mexico, Phytopathology, 87(10): 992-999
http://dx.doi.org/10.1094/PHYTO.1997.87.10.992
PMid:18945031
Hausbeck M.K., and Lamour K.H., 2004, Phytophthora capsici on vegetable crops: research progress and management challenges, Plant Disease, 88(12): 1292-1303
http://dx.doi.org/10.1094/PDIS.2004.88.12.1292
Ho H.H., Yu Y.N., Zhuang W.Y., and Liang Z.R., 1983, Mating types of heterothallic species of Phytophthora in China, Zhenjun Xuebao (Acta Mycologia Sinica), 2(3): 187-191 (何汉兴, 余永年, 庄文颖, 梁枝荣, 1983, 中国疫霉属异宗配合种的交配型, 真菌学报, 2(3): 187-191)
Islam S.Z., Babadoost M., Lambert K.N., Ndeme A., and Fouly H.M., 2005, Characterization of Phytophthora capsici isolates from processing pumpkin in Illinois, Plant Disease, 89(2): 191-197
http://dx.doi.org/10.1094/PD-89-0191
Jia J.S., 1992, On the chilli pepper Phytophthora blight and it’s control in Xinjiang, Zhiwu Bingli Xuebao (Acta Phytopathologica Sinica), 22(3): 257-262 (贾菊生, 1992, 新疆辣椒疫病及防控研究, 植物病理学报, 22(3): 257-262)
Lamour K.H., and Hausbeck M.K., 2001, Investigating the spatiotemporal genetic structure of Phytophthora capsici in michigan, Phytopathology, 91(10): 973-980
http://dx.doi.org/10.1094/PHYTO.2001.91.10.973
PMid:18944124
Li P., Chen Y.F., and Gao Z.M., 2011, Phylogenetic analysis among isolates of Phytophthora capsici in Anhui Province by RAPD, Anhui Nongye Daxue Xuebao (Journal of Anhui Agricultural University), 38(2): 287-291 (李萍, 陈艳芬, 高智谋, 2011, 安徽省辣椒疫霉菌株遗传多样性的RAPD分析, 安徽农业大学学报, 38(2): 287-291)
Liu Y.G., Zhang H.Y., Guo J.G., Lv H.P., He C.G., and Song S.Y., 2008, Mating type distribution and pathogenicity difference of phytophthora capsici in Gansu, Zhiwu Baohu Xuebao (Acta Phytophylacica Sinica), 35(5): 448-452 (刘永刚, 张海英, 郭建国, 吕和平, 贺春贵, 宋尚有, 2008, 甘肃省辣椒疫霉菌的交配型分布及其致病力差异, 植物保护学报, 35(5): 448-452)
Noon J.P., and Hickman C.J., 1974, Oospore production by a single isolate of Phytophthora capsici in the presence of chloroneb, Canadian Journal of Botany, 52(7): 1591-1595
http://dx.doi.org/10.1139/b74-208
Qi R.D., Wang T., Li P., Ding J.C., and Gao Z.M., 2012, Distribution of mating types of Phytophthora capsici and inheritance in asexual progenies in Anhui province, Zhiwu Bingli Xuebao (Acta Phytopathologica Sinica), 42(1): 45-50 (戚仁德, 汪涛, 李萍, 丁建成, 高智谋, 2012, 安徽省辣椒疫霉交配型的分布及在无性后代的遗传, 植物病理学报, 42(1): 45-50)
Silvar C., Merino F., and Díaz J., 2006, Diversity of Phytophthora capsici in Northwest Spain: Analysis of virulence, metalaxyl response, and molecular characterization, Plant Disease, 90(9): 1135-1142
http://dx.doi.org/10.1094/PD-90-1135
Wen J.Z., Jia W.X., Zhang M.H., and Yu W.Q., 1998, Species of Phytophthora causing stem and fruit rots of squash in Hei Longjiang province, Zhiwu Bingli Xuebao (Acta Phytopathologica Sinica), 28(3): 261-262 (文景芝, 贾文香, 张明厚, 于文全, 1998, 黑龙江省南瓜疫霉病病原菌鉴定, 植物病理学报, 28(3): 261-262)
Williams J.G.K., Kubelik A.R., Livak K.J., Rafalski J.A., and Tingey S.V., 1990, DNA polymorphisms amplified by arbitrary primers are useful as genetic markers, Nucleic Acids Res., 18(22): 6531-6535
http://dx.doi.org/10.1093/nar/18.22.6531
PMid:1979162 PMCid:332606
Yang M.Y., Cao J.F., Li X.D., Sun D.W., Wang C.Y., and Zhao Z.J., 2009, Molecular diagnosis and characterization of blight disease pathogen on pepper in Yunnan, Zhiwu Bingli Xuebao (Acta Phytopathologica sinica), 39(3): 297-303 (杨明英, 曹继芬, 李向东, 孙道旺, 王源超, 赵志坚, 2009, 云南辣椒疫病的分子诊断及其病原菌群体特征研究, 植物病理学报, 39(3): 297-303)
Yang X.H., 2004, Study on phytophthora blight of pepper in Guizhou province, Thesis for M.S., Southwest Agricultural University, Supervisor: Xiao C.G., pp.1-43 (杨学辉, 2004, 贵州省辣椒疫病研究, 硕士学位论文, 西南农业大学, 导师: 肖崇高, pp.1-43)
Zheng X.B., and Lu J.Y., 1990, Distribution of Phytophthora species and mating type in east and northeast China, Nanjing Nongye Daxue Xuebao (Journal of Nanjing Agricultural University), 13(3): 61-64 (郑小波, 陆家云, 1990, 华东、东北地区疫霉种的分布及交配型, 南京农业大学学报, 13(3): 61-64)