研究论文/Research Article
开发S5功能标记应用于水稻广亲和品种筛选 
2.中国农业科学院生物技术研究所, 北京, 100081
*同等贡献作者
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2014 年, 第 12 卷, 第 3 篇 doi: 10.5376/mpb.cn.2014.12.0003
收稿日期: 2014年01月23日 接受日期: 2014年01月24日 发表日期: 2014年03月23日
引用格式(中文):
吴爽等, 2014, 开发S5功能标记应用于水稻广亲和品种筛选, 分子植物育种(online), 12(3): 1016-1023 (doi: 10.5376/mpb.cn.2014.12.0003)
引用格式(英文):
Wu et al., 2014, Developing Functional Marker for S5 Gene and Screening Wide Compatibility Variety, Fenzi Zhiwu Yuzhong (online) (Molecular Plant Breeding), 12(3): 1016-1023 (doi: 10.5376/mpb.cn.2014.12.0003)
广亲和基因S5能有效地提高水稻亚种间杂交种的结实率,被广泛地应用于育种实践,如本单位以广亲和品种Cpslo17为S5基因供体育成的双优4183,已通过品种审定并应用于生产。本研究利用公开的S5分子信息,开发出基于PCR的功能性标记,对安徽省水稻地方品种等材料进行筛选,并对5份材料进行测序。研究结果表明:一份来源于东南亚材料含有S5基因,但332份安徽省水稻地方品种没有检测到S5基因,说明生殖隔离阻碍了基因交流,使得地方品种保留了遗传稳定性和多样性,同时也说明了全面收集和保存地方品种的重要性。
自20世纪60年代开始,随着全球人口的快速增长及农业用地被大量占用,粮食短缺问题引起了全球的广泛关注。为了满足对粮食的需求,到2025年粮食产量要在现有基础上(1995年全球谷物总产为1.68×1012 kg)增加50%以上(Khush, 2001)。水稻是最重要的粮食作物之一,要提高水稻总产量以保证粮食安全,只有依靠科技进步来提高单位面积产量。据统计,全球自20世纪60年代以来的40多年中,单产对水稻总产增长的贡献率达73% (朱德峰等, 2010)。水稻杂种优势利用对提高单产意义重大,可提高20%以上。基于形态性状、同工酶以及对一些化合物的不同反应,亚洲栽培稻(Oryza Sativa L.)分为籼(indica)、粳(japonica)2个亚种(Oka, 1974),籼亚种主要适应于热带和亚热带等低纬度地区,而粳亚种主要分布在高纬度和高海拔地区,从而形成天然的生殖隔离基因库。不同品种资源间杂种优势存在差异,籼粳间杂种优势明显高于品种间的杂种优势,主要表现在生物产量显著增加,根系发达,穗粒数增多。袁隆平认为在水稻单产突破中,利用亚种间杂种优势可能是在较短时期内取得成效的途径(袁隆平, 1987, 杂交水稻, (1): 1-3)。Zhang等(1997)利用9个籼稻和11个粳稻采用完全双列杂交设计,研究表明绝大多数亚种间F1的结实率低于50%,结实率的降低导致每穗实粒数减少从而影响最终产量的形成。广亲和种质的发现为解决亚种间杂交不亲和性提供可能,Ikehashi 和Araki(1984)从74个品种中筛选出Cpslo17等6个品种与籼、粳稻杂交F1都有较高的结实率,认为均具有广亲和特性且含有广亲和基因(wide compatibility gene, WCG)。卢诚和潘熙淦(1992)认为02428等的广亲和特性受1对主效基因控制,且与色素原基因C连锁。顾铭洪等(1993)通过广亲和品种之间的双列杂交分析得出Cpslo17、02428、轮回422等5个品种互为等位关系,受广亲和基因S5n控制,而Dular和Aus373则受另一对非等位基因控制,陈忠明等(1996)将来自Aus373的广亲和基因Sa(t)n初步定位于第7染色体上,其广亲和性比S5n弱。袁隆平等(1997)将水稻亚种间亲和性分为4大类,以热带粳稻(爪哇稻)如Cpslo17为第一类:广谱广亲和性,在育种上的实用性最好。Yanghihara等(1995)和Liu等(1997)利用不同群体分别找到与S5紧密连锁的位于第6染色体上的RFLP标记,之后该基因分别定位在40 kb、50 kb内,这2个定位结果存在着24.8 kb的重叠(Qiu et al., 2005; Ji et al., 2005)。Chen等(2008)成功地克隆了编码天冬氨酰蛋白酶的S5基因,它通过调控雌配子的育性来影响水稻结实率,亚种间杂种不育是由于籼粳等位基因间存在着两个碱基的差异导致了相应氨基酸的替换,而在广亲和品种,S5基因则有136 bp缺失导致功能丧失,所以与籼稻或粳稻杂交的F1结实率都表现正常。经过深入研究,发现在籼粳杂种的雌配子形成过程中,ORF4和ORF5基因共同作用杀死配子,籼型配子由于ORF3的保护而正常存活,而粳型配子因无保护而死亡,最终导致半不育,广亲和品种因无杀死配子的能力,故表现出广亲和特性(Yang et al., 2012)。本文就公开的分子信息开发出检测S5基因的功能性分子标记,希冀在安徽省水稻地方品种等材料中筛选到广亲和品种,为进一步配制亚种间组合或创制新的育种材料提供支撑。
1结果与分析
1.1功能性引物的实用性
为确定新设计引物的实用性,以含有广亲和S5基因的02428和Cpslo17与不含有S5的Nipponbare和9311进行PCR扩增(图1A)。由于广亲和品种02428和Cpslo17均有大片段的缺失,与Nipponbare和9311的带型存在明显差异,能较好地区分这两种类型的材料。
.png) 图1 S5功能性标记的电泳图 Figure 1 The PCR results of some rice varieties using S5 functional marker |
4183系本单位以Cpslo17为S5基因供体选育的粳型广亲和系(王守海, 2004, 作物研究, (4): 241),在S5这个位点上,与Cpslo17、培矮64S等广亲和材料的基因型完全相同(图1B),而且基因组序列也一致(图3)。1999年经扬州大学“863”项目广亲和系鉴定组鉴定,该品系程氏指数为22,属粳型,与4个广亲和性测验种(南特号、IR36、秋光、巴利拉)杂交F1的结实率分别为82.2%、86.5%、93.0%、90.7%,平均育性88.1%,广亲和性等级为优。以4183为父本育成的双优4183于2003年通过安徽省品种审定,表现结实率高、米质优,高产稳产。4183的育成再次证明了S5基因具有广谱广亲和性。
1.2 S5基因的筛查检测及部分品种S5位点序列比较
天鹅果是一个来自于东南亚的粳稻,用新设计的功能性引物S5cpslo筛查检测发现该材料与Cpslo17具有相同的带型(图2)。天鹅果穗粒数较多、农艺性状较好,其广亲和特性的鉴定为直接或间接利用该品种配制亚种间杂交品种提供了可能。
|
图2 功能标记S5 cpslo的筛查检测结果 Figure 2 The PCR result of screening rice landraces in Anhui province |
安徽水稻地方品种是各地农民在长期的生产实践中培育出来的地方良种,主要分布于皖南山区及江淮之间,淮河以北地区少有种植。在这些地方品种中绝大部分为籼稻,占97.6%,粳稻在江淮之间有少量种植。本研究利用功能性引物S5cpslo检测了332份安徽水稻地方品种,在所分析的安徽水稻地方品种中没有检测到S5基因(图2)。结果表明在进化过程中籼粳分化造就了丰富多样的稻种资源,但也导致生殖隔离,生殖隔离的存在阻碍了不同品种间的基因交流,使得这些地方品种保留了遗传稳定性和多样性,这能为发掘和利用蕴含的有利基因和分析水稻驯化过程中的选择事件提供实物。在二十世纪农家品种的整理收集过程中,部分品种可能被丢失,这也可能是没有检测到S5基因的一个原因。
在S5基因大片段缺失的上下游设计引物,用PCR产物直接测序,得图3。在S5这个位点上,Cpslo17、02428、天鹅果及4183的序列一致,都比非广亲和品种Balilla少了136 bp,且有2个相同的SNP(T→C,G→A)(其中T→C未显示)。表明S5基因的突变类型只有一种,即主要为大片段的缺失导致功能丧失,这与Chen 等(2008)的测序结果一致。
.png) 图3 5个品种在S5位点的序列比对 Figure 3 Sequence comparison of five varieties at S5 locus |
2讨论
2.1功能性标记的开发有助于提高育种效率
在育种过程中,选用合适的标记进行辅助选择,可减少盲目性,提高育种效率。遗传标记主要分为形态标记、生化标记和分子标记。广亲和基因S5与色素原基因C紧密连锁,水稻的紫稃尖色可作为筛选广亲和种质的标记,但紫稃尖性状是3对显性基因互作的结果,所以利用形态标记存在很大的误判导致实用性不强(Ikehashi and Araki, 1984)。由于所有广亲和品种都具有对醋酸-β-萘酯反应呈红色的酶带10A,余文金等提出用酯酶同工酶作为筛选广亲和种质的生化标记,但同工酶不能区分不等位的广亲和基因位点(余文金等, 1991, 遗传, (4): 5-8)。SSR、STS和CAPS等分子标记是以遗传物质为基础而开发的,具有可控性强、特异性高、鉴定快速准确、步骤简便,成为辅助育种的重要手段之一。邓启云等(2004)利用SSR引物进行分子标记辅助选择,将来自马来西亚普通野生稻高产位点yld1.1和yld2.1导入恢复系测64-7,育成新的优良恢复系远恢611。薛庆中等(1998)将抗白叶枯病的Xa21基因导入到水稻恢复系,准确率高达93.5%。张士陆等(2005)将糯性Wx基因导入到直链淀粉含量为28.4%的恢复系057中,使直链淀粉含量降至18%以下。方珊茹等(2013)将粒长主效基因GS3及其他外观品质性状基因导入到保持系Ⅱ-32B中,使得长宽比显著增大,而且垩白米率及垩白度极显著降低。本研究基于已克隆的S5基因的分子信息,通过选取广亲和品种和非广亲和品种进行测序,根据基因内部差异设计出分子标记,可有效地鉴定出广亲和品种,具有共显性、易检测、准确性高、实用性好等特点。
2.2广亲和基因S5在安徽省水稻地方品种中的分布
中国是亚洲栽培稻(Oryza sativa L.)原生起源地之一(郑殿升等, 2012),长江中下游是中国栽培稻的四个遗传多样性中心之一及可能的起源中心(黄燕红等, 2005),位于此区域的安徽省,水稻地方品种资源非常丰富,如耐肥、耐涝的大叶稻,耐瘠、耐旱的三粒寸,耐寒、耐旱、抗倒的顺水拖,还有塘稻(即深水稻)、宿县夹沟香稻等特异种质。在332份安徽省水稻地方品种中没有检测到S5基因,这表明在古老的地方品种间由于地理、生态、遗传基础等差异导致的生殖隔离,造就了遗传的多样性和丰富的稻种资源,同时也保留了地方品种在驯化过程中所形成的特有种性,这为利用和开发这些种质资源提供了有利条件;另一方面在20世纪七八十年代收集地方品种的过程中,可能遗失了一部分资源,使得本研究中所用的材料有所欠缺,这也警示我们全面收集古老地方品种的重要性。安徽水稻地方品种保留了米质优良、抗逆性强等一些性状,在育种上要利用这些优良性状配制亚种间组合,就可以通过多代回交结合分子标记辅助选择方法,将S5基因转育到亲本当中来解决亚种间结实率问题,S5cpslo标记的开发可使得育种目标能顺利实现。
栽培稻起源地区(含原生和次生起源地)存在着一定数量比较原始的、籼粳分化不彻底的类型,当中广亲和材料比较多,罗利军等总结了前人研究,认为广亲和基因可能有3个来源,而其中最多的是原始型的籼、粳稻(罗利军等, 1991, 作物品种资源, (1): 14-16),如云南省和东南亚山地的光壳陆稻、印度尼西亚的爪哇稻以及印度次大陆的Aus稻,在本研究中筛选出的广亲和种质天鹅果正是来源于东南亚。海拔跨度大的云南省是我国遗传多样性中心之一且是平均基因多样性最高的地方,长江中下游作为我国稻作的高产区,人工选择压大且稻作历史悠久,使得遗传多样性受到了削弱(黄燕红等, 2005),本实验结果也从某种程度上验证了这一点。至于安徽水稻地方品种中是否含有其他的广亲和基因有待于进一步研究。
2.3籼粳杂种优势应用的实践
亚种间杂种优势的利用一直是育种家追求的目标之一,培矮64S有来自爪哇稻Paddy血缘,是一个含有S5基因的籼型广亲和核不育系(罗孝和和袁隆平, 1989),以之为母本育成了培矮64S/C96211(王长义和方国成, 1999)、培矮64S/Hb-01(高金福等, 2000, 江西农业科技, (6): 8-10)、培两优2859(富昊伟等, 2006, 杂交水稻, 21(5): 79-80),其中培两优2859通过了浙江省品种审定,表现为分蘖较强、穗粒数多,结实率在80%以上。至于种子生产中,籼、粳花时差异在1 h以上而影响产量的问题,通过综合施用“调花宝”、“九二0”可有效地缓解花时不遇的负面影响。
赵志刚等(2010)利用分子标记辅助选择方法育成粳型亲籼不育系509S,该不育系聚合了S5、S7、S8和S9等4个育性相关位点,其中S7、S8和S9分别位于第7、第6、第4染色体上,表现为中性亲和性(Yanagihara et al., 1992; Wan et al., 1993; Wan et al., 1996)。509S与大部分籼稻杂交结实正常,其中表现突出的有亚种组合509S/W102。
马荣荣等(2010)以带有部分籼稻成分的粳型不育系甬粳2A,组配出甬优4号、6号、9号、11号、12号等亚种间组合,其中甬优6号、12号已被认定为超级杂交稻品种。用新设计的S5cpslo引物检测甬优12号的杂交F1种子,从图4中可以看出,4份甬优12号的杂交种在S5这个位点上都显现杂合状态,说明了在这个组合中至少存在一个广亲和基因,从而较好地解决了亚种间杂交种的结实率问题。
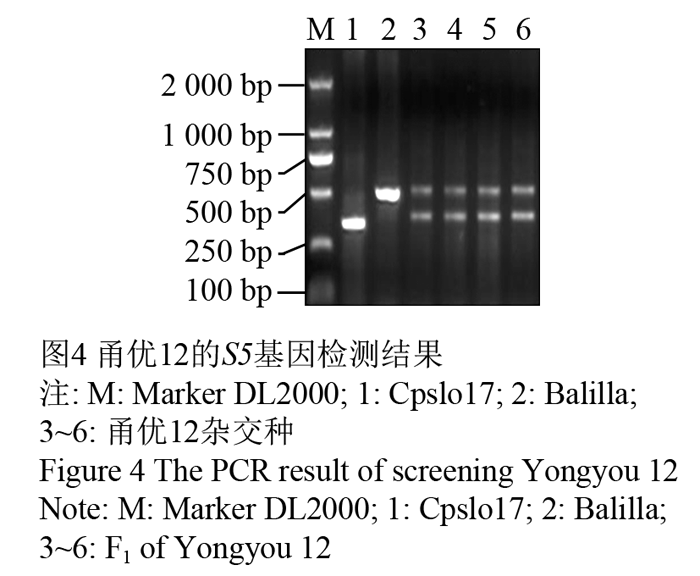 图4 甬优12的S5基因检测结果 Figure 4 The PCR result of screening Yongyou 12 |
3材料与方法
3.1试验材料
广亲和品种Cpslo17、02428、培矮64S和非广亲和品种Balilla、Nipponbare、9311等系从外单位引进,4183系本单位选育。供试材料332份安徽水稻地方品种(表1)由本单位收集与保存,天鹅果从外单位引进。
 表1 安徽水稻地方品种的来源地及数量 Table 1 The origin and number of rice landraces in Anhui province |
3.2广亲和基因S5功能标记设计及序列测定
根据已克隆的广亲和基因S5的分子信息,基于广亲和基因存在的136 bp的缺失,运用Primer 3在线设计引物(http://frodo.wi.mit.edu/),得到检测广亲和基因S5的特异性引物,S5cpslo-F:5'-CTGCACGTCTTGCT TCTTCA-3',S5cpslo-R:5'-AGCGAAGACAAAGGGAGTGA-3',在非广亲和品种中产物大小为492 bp,广亲和品种为356 bp。测序引物:S5Seq-F: 5'-ATGATCTTGCGGAAAACGAC-3',S5Seq-R:5'-GCTCGA TCGGATTAACAAGC-3',在非广亲和品种中产物大小为945 bp。获得的PCR产物直接测序,引物合成及测序均由上海生工生物工程技术服务有限公司完成。
3.3 DNA的提取及PCR反应程序
利用CTAB法提取水稻叶片的总DNA (Murray and Thompson, 1980),PCR反应总体系为20 μL,其中10×Buffer (MgCl2) 2.0 μL,2.5 mmol/L dNTP 1.0 μL,上下游引物各1.0 μL(10 μmol/L),Taq DNA聚合酶0.2 μL (5 U/μL),模板DNA 2.0 μL,ddH2O补足20 μL。PCR程序为94℃预变性5 min,94℃变性30 s,55℃退火30 s,72℃延伸60 s,30次循环,最后72℃延伸10 min。PCR产物加入指示剂在1.5%的琼脂糖凝胶中电泳30 min或直接用于测序。
3.4数据分析
5个品种的序列用DNAssit 2.2软件进行序列比对分析,“-”表示缺失,红色部分表示序列相同,本文中只显示部分序列。
作者贡献
吴爽和台德卫是本研究的实验设计和实验研究的执行人;吴爽完成数据分析,论文的写作;宛煜嵩、王德正参与实验研究并对论文进行修改;张效忠提供有关实验材料并参与实验设计,王守海全面指导实验设计,数据分析,论文的修改。全体作者都阅读并同意最终的文本。
致谢
本研究由安徽省农业科学院院长青年创新基金(11B10105)和国家高技术研究发展计划(2011AA10A100)共同资助。
参考文献
Chen J.J., Ding J.H., Ouyang Y.D., Du H.Y., Yang J.Y., Cheng K., Zhao J., Qiu S.Q., Zhang X.L., Yao J.L., Liu K.D., Wang L., Xu C.G., Li X.H., Xue Y.B., Xia M., Ji Q., Lu J.F., Xu M.L., and Zhang Q.F, 2008, A triallelic system of S5 is a major regulator of the reproductive barrier and compatibility of indica–japonica hybrids in rice, Proc. Natl. Acad. Sci. USA,105(32):11436-11441
http://dx.doi.org/10.1073/pnas.0804761105
PMid:18678896 PMCid:PMC2516230
Chen Z.M., Zou J.S., Zhu L.H., Li H.B., Gu F.L., and Gu M.H., 1996, Identification of RFLP markers linked with wide compatibility gene in variety Aus373 of ecotype Aus of rice by using bulked segregant analysis, Jiangsu Nongye Xuebao (Jiansu Journal of Agricultural Sciences), 12(4): 1-6 (陈忠明, 邹江石, 朱立煌, 李和标, 谷福林, 顾铭洪, 1996, 用分群分析法(BSA)鉴别与秋稻品种Aus373广亲和基因连锁的RFLP标记, 江苏农业学报, 12(4): 1-6)
Deng Q.Y., Yuan L.P., Liang F.S., Li J.M., Li X.Q., Wang L.G., and Wang B., 2004, Studies on yield-enhancing genes from wild rice and their marker-assisted selection in hybrid rice, Zajiao Shuidao (Hybrid Rice), 19(1): 6-10 (邓启云, 袁隆平, 梁凤山, 李继明, 李新奇, 王乐光, 王斌, 2004, 野生稻高产基因及其分子标记辅助育种研究, 杂交水稻, 19(1): 6-10)
Fang S.R., Wu C.Z., Liu Y.Q., Zheng P.L., Xiong X.J., Shen W.F., Zhuang L.P., and Zhao M.F., 2013, Molecular Marker-assisted Selection for Improving Appearance Quality of Ⅱ-32B, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 11(6):673-679 (方珊茹, 吴春珠, 刘玉芹, 郑苹立, 熊雪娇, 沈伟锋, 庄丽萍, 赵明富, 2013, 分子标记辅助选择改良Ⅱ-32B的外观品质, 分子植物育种, 11(6): 673-679)
Gu M.H., You A.Q., and Pan X.B., 1993, Genetic analysis on allelic relationship of wide compatibility genes among several WCV (Oryza sativa L.), Zhong guo Nongye Kexue (Scientia Agricultura Sinica), 26(1): 13-21 (顾铭洪,游艾青, 潘学彪, 1993, 水稻品种广亲和基因等位关系的遗传分析, 中国农业科学, 26(1): 13-21)
Huang Y.H., Sun X.L., and Wang X.K., 2005, Study on the center of genetic diversity and its origin of cultivated rice in China, Zhiwu Yichuan Ziyuan Xuebao (Journal of Plant Genetic Resources), 6(2): 125-129 (黄燕红, 孙新立, 王象坤, 2005, 中国栽培稻遗传多样性中心和起源研究, 植物遗传资源学报, 6(2): 125-129)
Ikehashi H., and Araki H., 1984, Varietal screening of compatibility type revealed in F1 fertility of distant crosses in rice, Jpn. J. Breed., 34: 304-313
http://dx.doi.org/10.1270/jsbbs1951.34.304
Ji Q., Lu J.F., Chao Q., Gu M.H., and Xu M.L., 2005, Delimiting a rice wide-compatibility gene S5n to a 50 kb region, Theor. Appl. Genet., 111(8): 1495-1503
http://dx.doi.org/10.1007/s00122-005-0078-0
PMid:16133303
Khush G.S., 2001, Challenges for meeting the global food and nutrient needs in the new millennium, Proc. Nutr. Soc., 60(1): 15-26
http://dx.doi.org/10.1079/PNS200075
PMid:11310421
Liu K.D., Wang J., Li H.B., Xu C.G., Liu A.M., Li X.H., and Zhang Q.F., 1997, A genome-wide analysis of wide compatibility in rice and the precise location of the S5 locus in the molecular map, Theor. Appl. Genet., 95(5-6): 809-814
http://dx.doi.org/10.1007/s001220050629
Lu C., and Pan X.G., 1992, Inheritance of wide compatibility in rice cultivars 02428 and 8504, Zhongguo Shuidao Kexue (Chinese Journal of Rice Science), 6(3): 113-118 (卢诚, 潘熙淦, 1992, 水稻品种02428和8504的广亲和性遗传, 中国水稻科学, 6(3): 113-118)
Luo X.H., and Yuan L.P., 1989, Breeding of rice wide compatibility lines, Zajiao Shuidao (Hybrid Rice), (2): 35-38 (罗孝和, 袁隆平, 1989, 水稻广亲和系的选育, 杂交水稻, (2): 35-38)
Ma R.R., Wang X.Y., Lu Y.F., Zhou H.C., Cai K.F., Li X.H., and Zhang Z.Y., 2010, Breeding and application of late japonica CMS line Yongjing 2A and its late indica-japonica hybrid rice Combinations, Zajiao Shuidao (Hybrid Rice), 25(S): 185-189 (马荣荣, 王晓燕, 陆永法, 周华成, 蔡克锋, 李信华, 章志远, 2010, 晚粳不育系甬粳2号A及其籼粳杂交晚稻组合的选育及应用, 杂交水稻, 25(S): 185-189)
Murray M.G., and Thompson W.F., 1980, Rapid isolation of high molecular weight plant DNA, Nucleic. Acids. Res., 8(19): 4321-4325
http://dx.doi.org/10.1093/nar/8.19.4321
PMid:7433111 PMCid:PMC324241
Oka H.I., 1974, Experimental studies on the origin of cultivated rice, Genetics, 78(1): 475-486
PMid:17248664 PMCid:PMC1213206
Qiu S.Q., Liu K.D., Jiang J.X., Song X., Xu C.G., Li X.H., and Zhang Q.F., 2005, Delimitation of the rice wide compatibility gene S5n to a 40-kb DNA fragment, Theor. Appl. Genet., 111(6): 1080-1086
http://dx.doi.org/10.1007/s00122-005-0033-0
PMid:16177904
Wan J.M., Yamaguchi Y., Kato H., and Ikehashi H., 1996, Two new loci for hybrid sterility in cultivated rice (Oryza sativa L.), Theor. Appl. Genet., 92(2): 183-190
http://dx.doi.org/10.1007/BF00223375
PMid:24166167
Wan J.M., Yanagihara S., Kato H., and Ikehashi H., 1993, Multiple alleles at a new locus causing hybrid sterility between a Korean indica variety and japonica variety in rice, Jpn. J. Breed., 43: 507-516
http://dx.doi.org/10.1270/jsbbs1951.43.507
Wang C.Y., and Fang G.C., 1999, Development of new inter-subspecies combination Peiai64S/C96211 in two-line hybrid rice, Hubei Nongye Kexue (Hubei Agricultural Sciences), (2): 3-5 (王长义, 方国成, 1999, 两系法亚种间杂交稻新组合培矮64S/C96211, 湖北农业科学, (2): 3-5)
Xue Q.Z., Zhang N.Y., Xiong Z.F., Li C., and Zhu L.H., 1998, The development of the rice restorer lines with the resistance of the bacterial blight disease by the marker assisted selection, Zhejiang Nongye Daxue Xuebao (Journal of Zhejiang Agricultural University), 24(6): 581-582 (薛庆中, 张能义, 熊兆飞, 李翀, 朱立煌, 1998, 应用分子标记辅助选择培育抗白叶枯病水稻恢复系, 浙江农业大学学报, 24(6): 581-582)
Yanagihara S., Kato H., and Ikehashi H., 1992, A new locus for multiple alleles causing hybrid sterility between an Aus variety and javanica varieties in rice (Oryza sativa L.), Jpn. J. Breed., 42(4): 793-801
http://dx.doi.org/10.1270/jsbbs1951.42.793
Yang J.Y., Zhao X.B., Cheng K., Du H.Y., Ouyang Y.D., Chen J.J., Qiu S.Q., Huang J.Y., Jiang Y.H., Jiang L.W., Ding J.H., Wang J., Xu C.G., Li X.H., and Zhang Q.F., 2012, A killer-protector system regulates both hybrid sterility and segregation distortion in rice, Science, 337(6100): 1336-1340
http://dx.doi.org/10.1126/science.1223702
PMid:22984070
Yuan L.P., Wu X.J., Yan Y.C., and Luo X.H.,1997, A strategy for developing wide spectrum compatibility rice line, Zhongguo Nongye Kexue (Scientia Agricultrua Sinica), 30(4): 1-8 (袁隆平, 武小金, 颜应成, 罗孝和, 1997, 水稻广谱广亲和系的选育策略, 中国农业科学, 30(4): 1-8)
Zhang Q.F., Liu K.D,, Yang G.P., Saghai Maroof M.A., Xu C.G., and Zhou Z.Q.,1997, Molecularmarker diversity and hybrid sterility in indica-japonica rice crosses, Theor. Appl. Genet., 95(1-2): 112-1181
http://dx.doi.org/10.1007/s001220050538
Zhang S.L., Ni D.H., Yi C.X., Li L., Wang X.F., Wang Z.Y., and Yang J.B., 2005, Reducing amylose content of indica rice variety 057 by molecular marker-assisted selection, Zhongguo Shuidao Kexue (Chinese Journal of Rice Science), 19(5): 467-470 (张士陆, 倪大虎, 易成新, 李莉, 汪秀峰, 王宗阳, 杨剑波, 2005, 分子标记辅助选择降低籼稻057的直链淀粉含量, 中国水稻科学, 19(5): 467-470)
Zhao Z.G., Chen L.M., Jiang L., and Wan J.M., 2010, Breeding of an indica-compatible japonica photoperiod-sensitive genic male sterile line in rice and its heterosis, Zajiao Shuidao (Hybrid Rice), 25(S): 256-263 (赵志刚, 陈亮明, 江玲, 万建民, 2010, 水稻粳型亲籼光温敏不育系509S的选育及杂种优势, 杂交水稻, 25(S): 256-263)
Zheng D.S., Liu X., and Li Y., 2012, Cultivated plants originated in China, Zhiwu Yichuan Ziyuan Xuebao (Journal of Plant Genetic Resources), 13(1): 1-10 (郑殿升, 刘旭, 黎裕, 2012, 起源于中国的栽培植物, 植物遗传资源学报, 13(1): 1-10)
Zhu D.F., Cheng S.H., Zhang Y.P., Lin X.Q., and Chen H.Z., 2010, Analysis of Status and Constraints of Rice Production in the World, Zhongguo Nongye Kexue (Scientia Agricultura Sinica), 43(3): 474-479 (朱德峰, 程式华, 张玉屏, 林贤青, 陈惠哲, 2010, 全球水稻生产现状与制约因素分析, 中国农业科学, 43(3): 474-479)