转blti2抗寒基因棉花的抗寒性分析 
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2011 年, 第 9 卷, 第 88 篇 doi: 10.5376/mpb.cn.2011.09.0088
收稿日期: 2011年05月19日 接受日期: 2011年06月27日 发表日期: 2011年07月05日
王冬梅等, 2011, 转blti2抗寒基因棉花的抗寒性分析, 分子植物育种 Vol.9 No.88 (doi: 10.5376/mpb.cn.2011.09.0088)
来自大麦的blti2 (barley low temperature-induced)低温诱导基因,是否能在其他作物中表现出抗性尚没有报道。本文利用花粉管通道法将其转入棉花中并得到了纯合系。通过RT-PCR的检测,blti2在转基因棉中稳定遗传;在4℃低温下处理不同时间测定可溶性糖、脯氨酸和丙二醛的含量,结果发现,与对照相比,可溶性糖和脯氨酸随处理时间延长呈现快速增长的趋势,与抗寒性呈正相关,而丙二醛则呈现下降趋势,与抗寒性呈负相关;在-4℃低温下处理不同时间,转基因棉可以持续6 h,对照全部倒伏而死。这些结果表明转blti2基因棉花的抗寒性得到了提高,为创建棉花抗寒种质资源奠定了基础。
低温胁迫是植物栽培中常常遇到的一种灾害,由于低温在植物整个生育过程中均会造成不利影响,如冷害会造成植株苗弱、生长迟缓、萎蔫、黄化、局部坏死、产量降低和品质下降等不良影响,对农业生产造成严重损失。同样,在新疆北部大部分棉田,每年都会受到倒春寒不同程度的影响,造成棉花大面积的烂种、死苗致使补种、重播,增加了生产成本。因此,创建抗寒棉花种质资源对培育新的棉花品种具有十分重要的作用。
近年来,利用遗传转化技术将外源抗寒基因转入不同作物中取得了很大进展,例如,来自胡萝卜36KD抗冻蛋白(徐文丽, 2005; 魏宪辉, 2007)、拟南芥CBF3和COR15a (吴纯清, 2006)、沙冬青AmLEA和AmPI基因(陈奕吟, 2007)、小麦抗寒基因CBF (Shigo et al, 2007, Plant Physiology and Biochemistry, doi:10.1016/j.plaphy.2007.10.019)和昆虫的抗冻蛋白(王艳等, 2008)等转入烟草,拟南芥CBF1转入水稻、地被石竹和草莓(金建凤等, 2005; 吴琰等, 2007; 金万梅等, 2007),拟南芥CBF4转入辣椒(王兴娥等, 2008),这些转基因作物经过低温试验或生理生化指标测定,均证明外源抗寒基因提高了作物的抗寒性。
Bahn等(2001)利用SSH (Suppression Subtractive Hybridization)方法获得了160个cDNA blti (barley low temperature-induced),其中blti2是ABA (Abscisic acid)非依赖途径的低温诱导基因,它编码164个氨基酸,通过比对,与大麦中的低温应答(Low Temperature-Responsive, LTR)基因无同源性,是低温处理下的穿膜基因。将blti2基因转入棉花中尚未见报道,本文利用RT-PCR法获得了blti2基因,通过花粉管通道法将其转入棉花中,通过分子检测、南繁加代和自交,已经得到T4代株系,经过低温试验后,对其中一个抗寒性表现好的株系进行了相同低温下不同处理时间的生理指标测试,来研究转基因棉的抗寒性,也为创建棉花抗寒种资资源奠定基础。
1结果与分析
1.1转基因棉中blti2基因的遗传稳定性分析
采集转blti2基因棉花株系的几个抗卡那霉素植株,提取总RNA,反转录后,用特异引物检测blti2基因,在随机采集的抗卡那霉素植株中均有blti2基因的存在,表明blti2基因稳定遗传(图1)。
图1 转基因棉的RT-PCR检测 Figure 1 RT-PCR testing of the transgenic cotton |
1.2脯氨酸含量变化规律分析
根据脯氨酸不同浓度下的吸光值计算出回归方程式,得到不同样品在低温下不同处理时间后脯氨酸含量的变化趋势,转基因棉在1 d时,其脯氨酸含量为0.268 μg/g低于对照,随后快速上升,到第5 d时达到峰值,为0.571 μg/g,超过对照20.97%,接着下降。而对照缓慢上升,其含量均低于转基因棉,到5 d时开始下降,含量为0.472 μg/g,然后与转基因棉的脯氨酸含量慢慢接近(图2)。
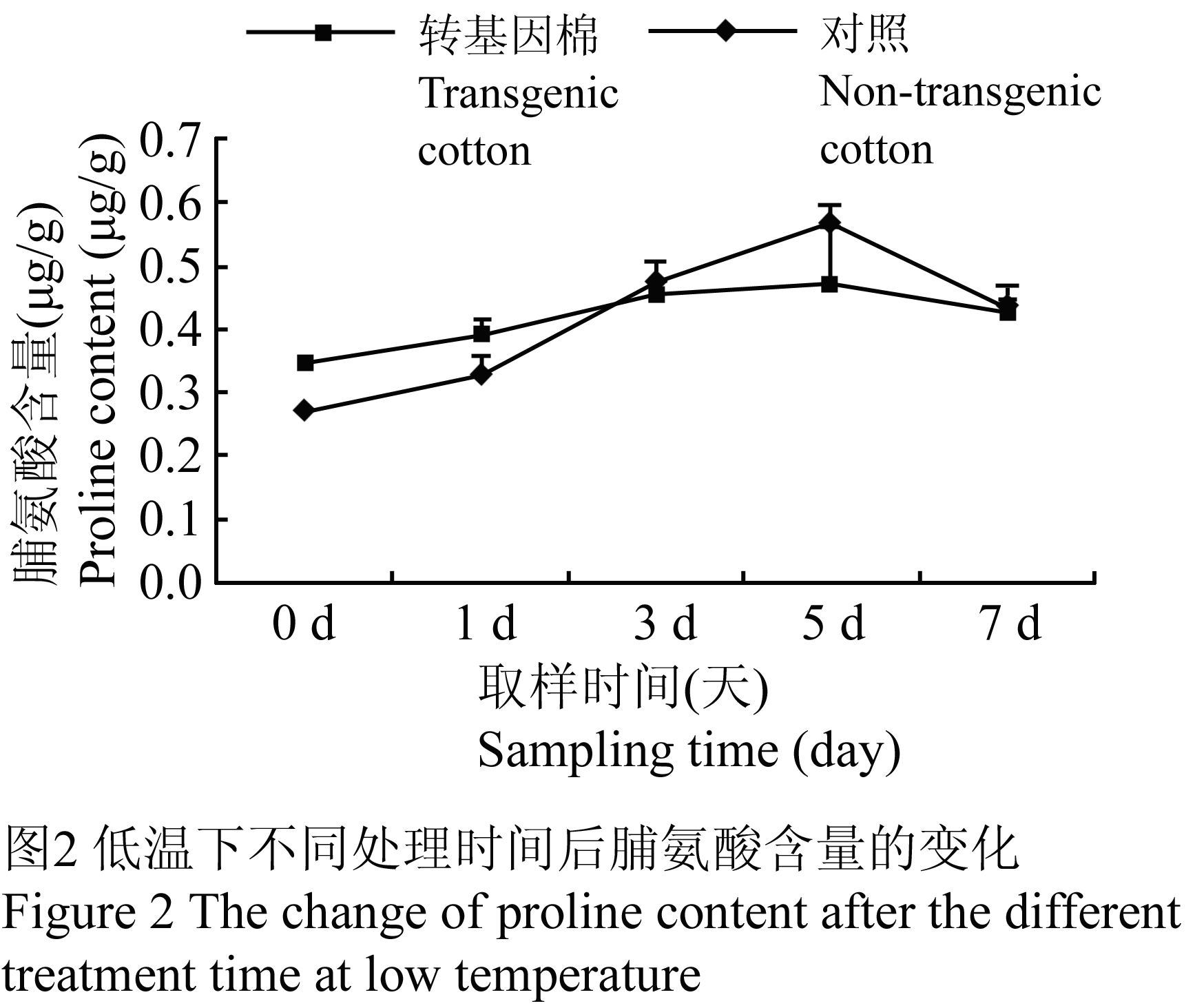 Figure 2 The change of proline content after the different treatment time at low temperature |
1.3可溶性糖含量变化规律分析
通过回归方程计算出不同样品在低温下不同处理时间后可溶性糖含量的变化趋势,从(图3)可以看出,在低温处理1 d时,转基因棉和对照可溶性糖含量接近,随后,转基因棉快速上升到第5 d达到峰值,为279.233 μg/g,超过对照33.95%。而对照在第3 d达到峰值,为217.224 μg/g,随后,转基因棉和对照都开始下降,在第7 d天时,分别为227.876 μg/g和143.751 μg/g(图3)。
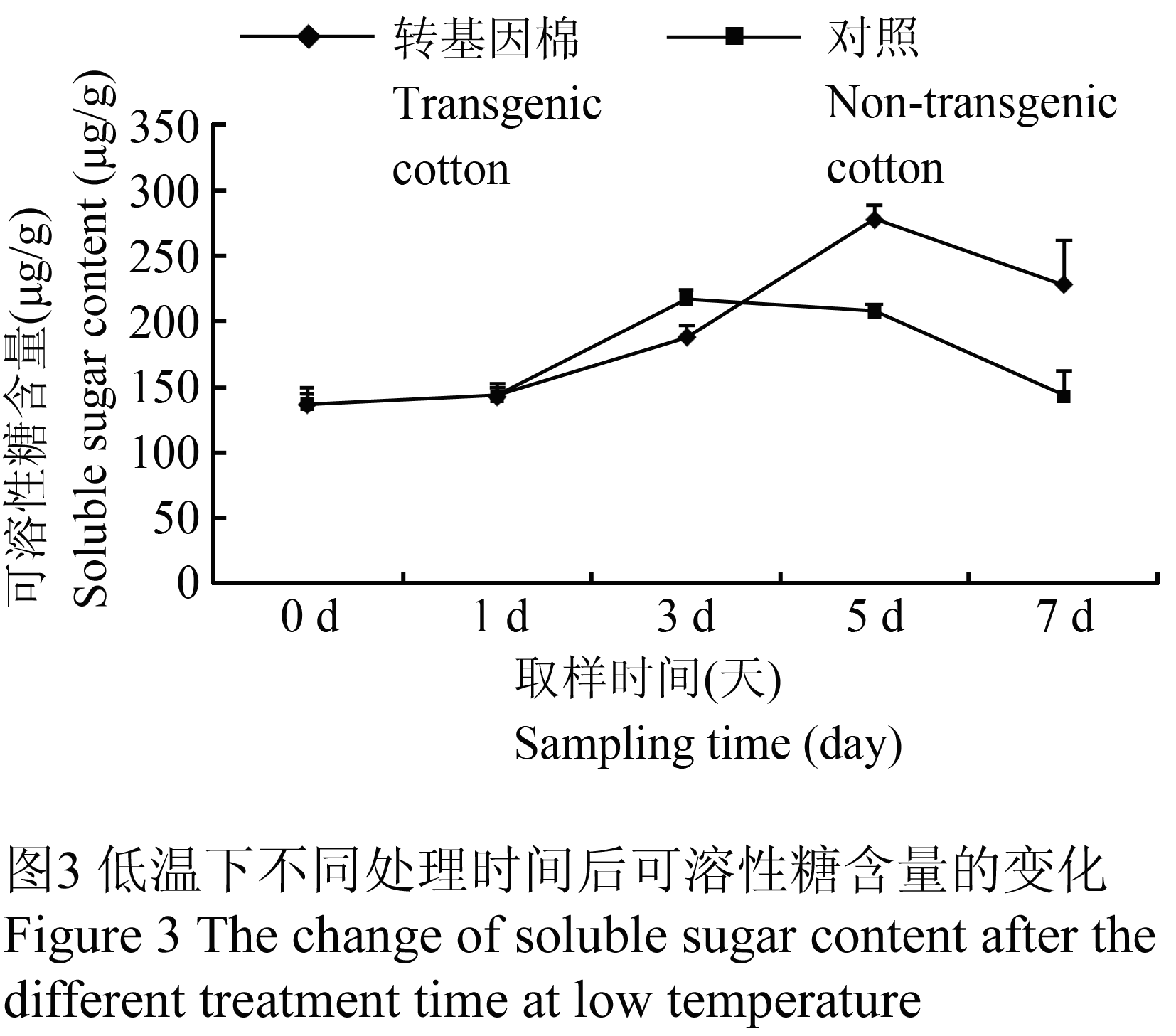 Figure 3 The change of soluble sugar content after the different treatment time at low temperature |
1.4丙二醛(MDA)含量变化规律分析
利用双组分分光光度计法建立方程组,根据植物组织的重量计算测定样品中MDA的含量,从(图4)可以看出,未经过低温处理的转基因棉的MDA含量明显高于对照,低温处理后快速下降,到1 d时,达到最低点,为15.992 μmoL/g,低于对照28.33%,而对照的MDA含量在1 d时无明显变化。随后,尽管转基因棉和对照的变化相同,呈现上升趋势,在第3 d时达到峰值,分别为33.356 μmoL/g和38.386 μmoL/g,然后接着下降,但是转基因棉的MDA含量始终低于对照,在第7 d时分别为16.376 μmoL/g和24.331 μmoL/g,低于对照48.58% (图4)。
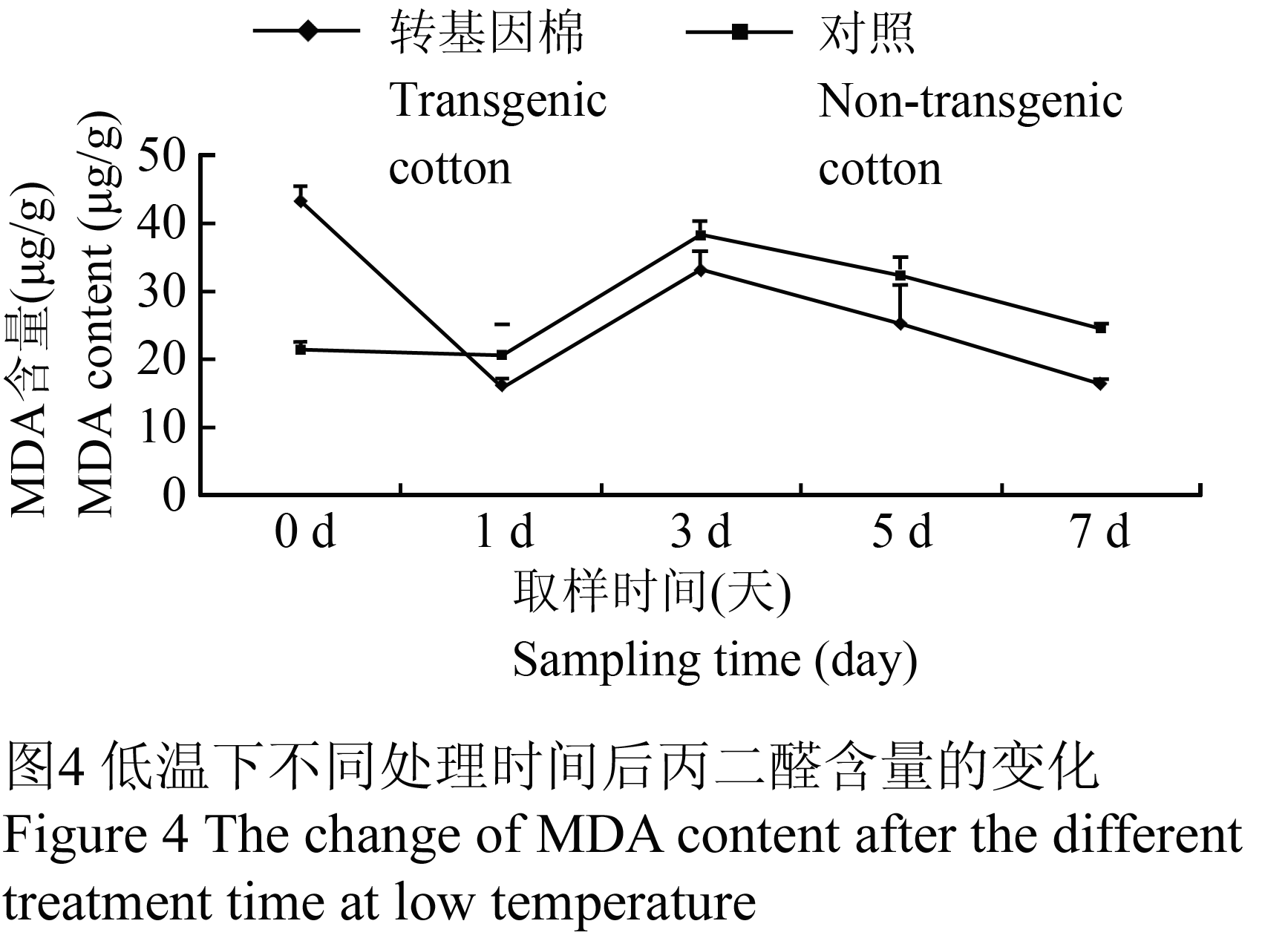 图4 低温下不同处理时间后丙二醛含量的变化 Figure 4 The change of MDA content after the different treatment time at low temperature |
1.5低温耐受性分析
将转基因棉和对照同时放入-4℃的培养箱中,1 h观察1次,在2 h时,转基因棉和对照都无变化,3 h时,对照叶边缘开始出现水渍状,4 h时,59.38%的对照叶片出现水渍状,呈冻伤状态,5 h时,有90.63%的对照叶片大面积出现水渍状,但尚未倒伏,而个别转基因棉叶边缘开始出现水渍状,到6 h有97.92%的对照全部倒伏,14.02%的转基因棉叶片大面积出现水渍化,8 h时,有36.45%的转基因棉开始倒伏,而倒伏植株叶片并未呈水渍化,9 h时,有77.57%转基因棉倒伏,叶片仍保持原状(图5)。
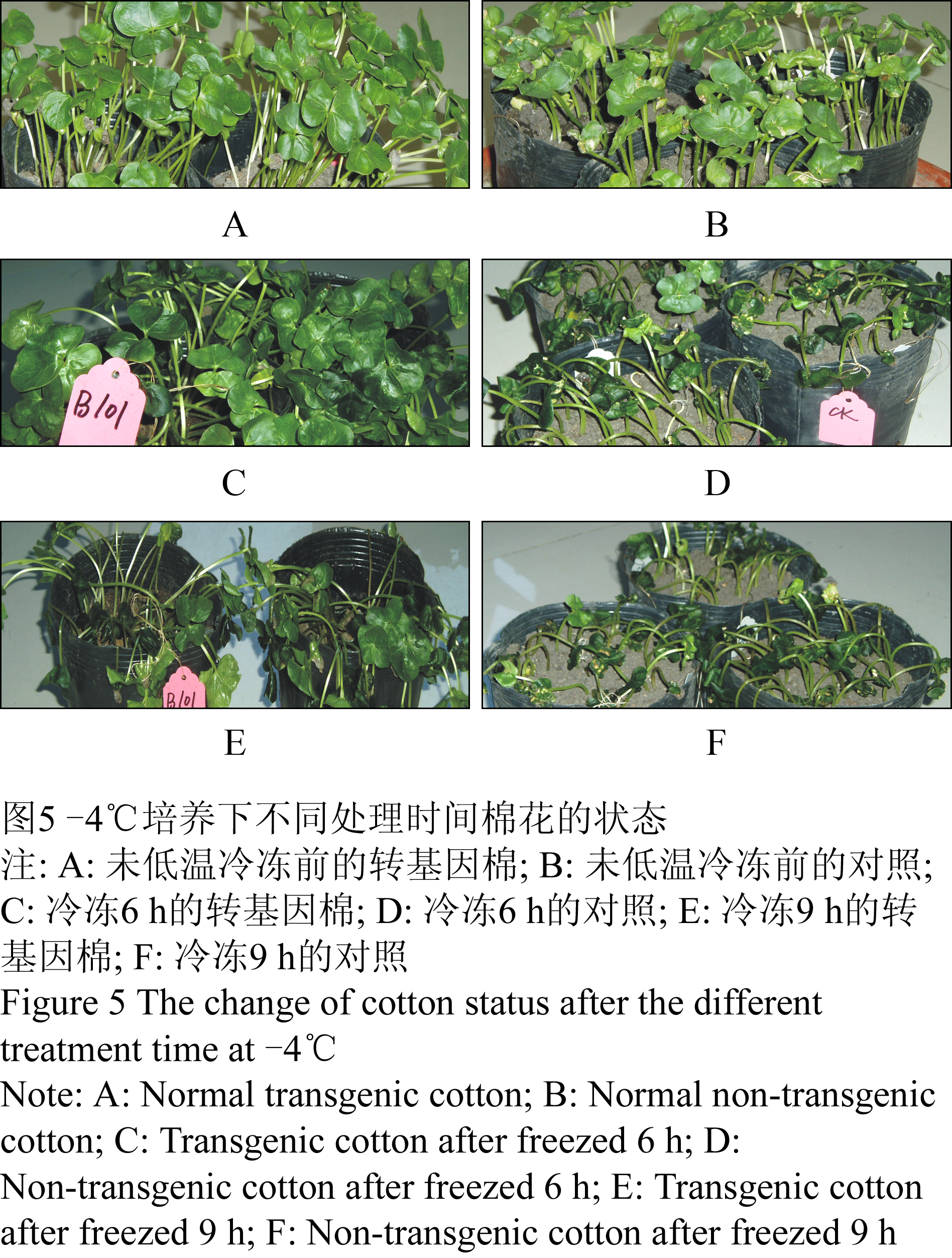 图5 -4℃培养下不同处理时间棉花的状态 Figure 5 The change of cotton status after the different treatment time at -4℃ |
2讨论
关于植物抗寒性测定的生化指标多有研究,其中脯氨酸、可溶性糖和丙二醛(MDA)等指标应用最多,大多数学者认为脯氨酸作为一种重要的渗透调节物质,在逆境胁迫下,对植物的代谢具有调节作用,脯氨酸含量与植物抗寒性呈正相关。可溶性糖是植物体内最重要的渗透调节物质和能量物质,可以降低细胞的结冰温度,减轻冻害,其含量与植物抗寒性呈正相关。MDA是膜脂过氧化的产物之一,可以反映细胞膜脂过氧化程度及细胞遭受伤害的程度,在低温胁迫下,植物体内活性氧自由基的积累超出一定限度时,就会引起膜脂过氧化,MDA产物大量积累,其含量与植物抗寒性呈负相关(陶雅和孙启忠, 2008; 张成军等, 2008; 颜范悦等, 2009; 高京草等, 2010; 司剑华和卢素锦, 2010; 张保青等, 2011)。也有一些学者持不同的观点,例如(龚双军等, 2005, 中国棉花, 32(3): 16-17)在研究棉花品种苗期抗寒性及其生理指标测定时,即认为供试8个材料的幼苗对低温的抗性有明显区别,其可溶性糖含量和丙二醛含量的提高幅度与品种抗寒性呈明显的正相关,何开跃等(2004)认为,可溶性糖与MDA指标可作为含笑抗寒性生理指标,用于含笑及种源抗寒性选择,脯氨酸与含笑树种间抗寒性的关系不明显。
本文选用脯氨酸、可溶性糖和丙二醛等3个生化指标来研究转blti2抗寒基因棉花的抗寒性,将转基因棉和受体对照同时放入4℃培养箱,和对照相比,转基因棉的脯氨酸含量和可溶性糖含量都超过对照,表明转基因棉的抗寒性与脯氨酸和可溶性糖含量呈正相关;而转基因棉的丙二醛含量一直低于对照,其抗寒性与丙二醛含量呈负相关,这与大多数学者持相同观点。此外,将转基因棉和受体对照同时经过-4℃的冷冻处理,相比对照,转基因棉对冷冻的耐受力可以持续6 h,若从降低产量损失来看,则可以持续8 h。以上数据和现象都说明经过前期的低温诱导,抗寒基因逐渐启动,调控低温保护物质开始增加,如脯氨酸和可溶性糖增加,其叶片细胞内的渗透势和水势降低,增强了细胞渗透调节能力,同时细胞内活性氧自由基减少,控制了细胞内膜脂过氧化,减小了低温对膜脂的损害,MDA积累降低,致使转抗寒基因棉花比对照增加了抵抗低温的能力,提高了转抗寒基因棉花的抗寒性。
总之,通过本文的研究,来自大麦的blti2基因可以使棉花的抗寒性有一定的提高,这为创建棉花抗寒种质资源奠定了较好的基础。随后,我们将对低温下转基因棉的膜结构、相关保护酶的变化以及蛋白表达等做进一步研究,以对植物的抗寒机理作有益探索。
3材料与方法
3.1材料
通过花粉管通道法获得转blti2抗寒基因棉花T4代株系B101,由新疆农业科学院核技术生物技术研究所农作物生物技术重点实验室提供。
3.2方法
3.2.1转基因棉中blti2基因的遗传稳定性检测
采集幼嫩叶片迅速冻于液氮中,提取棉总RNA,提取过程按刘洋等(2006)的方法。利用Promega的反转录体系进行blti2基因的RT-PCR检测。
3.2.2待测材料的低温处理
将转基因棉花和对应的非转基因棉对照(新陆早23号)播种于花盆中,待幼苗长到2~3片完全展开的真叶时,放入4℃培养箱中,分别于0 d、1 d、3 d、5 d、7 d采样,每处理重复三次。
3.2.3脯氨酸含量测定
准确称取经低温处理的不同时间的待测棉花叶片各0.3 g,利用茚三酮显色法测定脯氨酸含量,方法详见(邹琦, 2000, 中国农业出版社, 161-162)。
3.2.4可溶性糖含量测定
准确称取经低温处理的不同时间的待测棉花叶片各0.3 g,利用蒽酮一硫酸显色法测定可溶性糖含量,方法详见(邹琦, 2000, 中国农业出版社, 111-112)。
3.2.5丙二醛(MDA)含量测定
准确称取经低温处理的不同时间的待测棉花叶片各0.3 g,利用硫代巴比妥酸法测定MDA含量,方法详见(邹琦, 2000, 中国农业出版社, 173-174)。
3.2.6低温耐受性试验
将转基因棉和受体对照同时放在-4℃培养箱内,每1 h观察转基因棉和对照的受冻情况,直至转基因棉和对照全部冻死为止。
作者贡献
王冬梅、李晓荣和李静是本研究的实验设计和实验研究的执行人;李静完成数据分析;张帅参与实验设计,试验结果分析;王冬梅是项目的构思着和负责人,指导实验设计,数据分析,论文写作与修改;黄乐平是项目的构思者。全体作者都阅读并同意最终的文本。
致谢
本研究由新疆维吾尔自治区重大专项(200731138-3)资助。作者感谢新疆农业科学院核技术生物技术研究所高孟萍女士在本实验过程中的帮助。感谢两位匿名的同行评审人的评审建议和修改建议。
参考文献
Bahn S.C., Bae M.S., Park Y.B., Oh S.I., Jeung J.U., Bae J.M., Chung Y.S., and Shin J.S., 2001, Molecular cloning and characterization of a novel low temperature-induced gene, blti2, from barley (Hordeum vulgare L.), Biochimica Biophysica Acta, 1522(2): 134-137
Chen Y.Y., 2007, Expression of the cold-induced Genes of Ammopiptanthus mongolicus in tobacco and the cold tolerance of the transgenetic plants, Thesis for M.S., Beijing Forestry University, Supervisor: Lu C.F., pp.1-68 (陈奕吟, 2007, 沙冬青低温诱导基因转化烟草及转基因植物的抗寒性研究, 硕士学位论文, 北京林业大学, 导师: 卢存福, pp.1-68)
Gao J.C., Wang H.X., and Li X.X., 2010, Relationship between Soluble protein, MDA, and jujube tree cold hardiness, Beifang Yuanyi (Northern Horticulture), 23: 18-20 (高京草, 王慧霞, 李西选, 2010, 可溶性蛋白, 丙二醛含量与枣树枝条抗寒性的关系研究, 北方园艺, 23: 18-20)
He K.Y., Li X.C., Huang L.B., and Sun Y.Z., 2004, A study on cold-resistant mechanism in three kinds of michelia, Nanjing Linye Daxue Xuebao (Journal of Nanjing Forestry University (Natural Sciences Edition)), 28(4): 62-84 (何开跃, 李晓储, 黄利斌, 孙玉珍, 2004, 3种含笑耐寒生理机制研究, 南京林业大学学报(自然科学版), 28(4): 62-64)
Jin J.F., Gao Q., Chen Y., and Wang J.H., 2005, Transfer of Arabidopsis CBF1 gene leads to increased praline contents in rice plant, Xibao Shengwuxue Zazhi (Chinese Journal of Cell Biology), 27: 73-76 (金建凤, 高强, 陈勇, 王君晖, 2005, 转移拟南芥CBF1基因引起水稻植株脯氨酸含量提高, 细胞生物学杂志, 27: 73-76)
Jin W.M., Dong J., Yin S.P., Yan A.L., and Chen M.X., 2007, CBF1 gene transgenic strawberry and increase freezing tolerance, Xibei Zhiwu Xuebao (Acta Botanica Boreali-Occidentalia Sinica), 27(2): 223-227 (金万梅, 董静, 尹淑萍, 闫爱玲, 陈梅香, 2007, 冷诱导转录因子CBF1转化草莓及其抗寒性鉴定, 西北植物学报, 27(2): 223-227)
Liu Y., He X.R., Ma H.B., Wu Y.L., and Yang Y.M., 2006, Extraction of total RNA from cotton (Gossypium hirsutum L.) tissues with CTAB-PVP method, Zhongguo Nongye Daxue Xuebao (Journal of China Agriculture University), 11(1): 53-56 (刘洋, 何心娆, 马红波, 吴泳历, 杨佑明, 2006, 用CTAB-PVP法提取棉花各组织总RNA的研究, 中国农业大学学报, 11(1): 53-56)
Si J.H., and Lu S.J., 2010, Effects of low temperature stress on cold-resistance physiologicalindexes of five Tamarix L. in Qinghai, Zhongnan Linye Keji Daxue Xuebao (Journal of Central South University of Forestry& Technology), 30(8): 78-81 (司剑华, 卢素锦, 2010, 低温胁迫对5种柽柳抗寒性生理指标的影响, 中南林业科技大学学报, 30(8): 78-81)
Tao Y., and Sun Q.Z., 2008, Dynamic variation of soluble sugar , tota l nitrogen and malondialdehyde in different alfalfa varieties and their effect on alfalfa’s cold resistance, Zhongguo Nongye Keji Daobao (Journal of Agricultural Science and Technology), 10(S1): 56-60 (陶雅, 孙启忠, 2008, 不同紫花苜蓿品种可溶性糖, 全氮, 丙二醛含量动态变化及其与抗寒性关系研究, 中国农业科技导报, 10(S1): 56-60)
Wang X.E., Gong Z.H., Li D.W., Chen R.G., Lu M.H., and Huang W., 2009, Transformation of pepper with cold-induced gene c-repeat binding factor 4 (CBF4) and analysis of cold hardness-related indexe, Nongye Shengwu Jishu Xuebao (Journal of Agricultural Biotechnolog), 17(5): 830-835 (王兴娥, 巩振辉, 李大伟, 陈儒钢, 逯明辉, 黄炜, 2009, 冷诱导基因C-重复基序结合因子4 (CBF4)在辣椒中的遗传转化及抗寒性分析, 农业生物技术学报, 17(5): 830-835)
Wang Y., Qiu L.M., Xie W.J., Huang W., Ye F., Zhang F.C., and Ma J., 2008, Cold tolerance of transgenic tobacco carrying gene encoding insect antifreeze protein, Zuowu Xuebao (Acta Agronomica Sinica), 34(3): 397-402 (王艳, 邱立明, 谢文娟, 黄薇, 叶锋, 张富春, 马纪, 2008, 昆虫抗冻蛋白基因转化烟草的抗寒性, 作物学报, 34(3): 397-402)
Wei X.H., 2007, Cold tolerance and the genetic characteristics of tobacco plants transformed with a carrot 36KD antifreeze protein gene, Thesis for M.S., Beijing Forestry University, Supervisor: Lu C.F., pp.1-60 (魏宪辉, 2007, 转抗冻蛋白基因烟草的抗寒性及遗传稳定性分析, 硕士学位论文, 北京林业大学, 导师: 卢存福, pp.1-60)
Wu C.Q., 2006, Influence of CBF3 and COR15a on Cold resistance of tobacco, Thesis for M.S., Xinan University, Supervisor: LIang G.L., and Zhang X.G., pp.1-50 (吴纯清, 2006, CBF3和COR15a基因对烟草抗寒性的研究, 硕士学位论文, 西南大学, 导师: 梁国鲁, 张兴国, pp.1-50)
Wu Y., Dong J., Guo B.L., Lu R.Q., and Jin W.M., 2007, Estimating cold-resistance of transgenic Dianthus chinese with CBF1 Gene, Zhongguo Nongxue Tongbao (Chinese Agricultural Science Bulletin), 23(5): 59-62 (吴琰, 董静, 郭宝林, 鲁韧强, 金万梅, 2007, 转CBF1基因地被石竹的抗寒性评价, 中国农学通报, 23(5): 59-62)
Xu W.L., 2005, Expression of a carrot 36KD antifreeze protein gene in tobacco and the cold tolerance of the transgenic plants, Thesis for M.S., Beijing Forestry University, Supervisor: Lu C.F., pp.1-57 (徐文丽, 2005, 抗冻蛋白基因转化烟草及转基因植株抗寒性研究, 硕士学位论文, 北京林业大学, 导师: 卢存福, pp.1-57)
Yan F.Y., Jiang N., Pei X.H., and Zhu P.F., 2009, Research for the Chill-resistance of Bulb in Lily, Beifang Yuanyi (Northern Horticulture), 9: 154-157 (颜范悦, 姜楠, 裴新辉, 祝朋芳, 2009, 百合种球抗寒性的研究, 北方园艺, 9: 154-157)
Zhang B.Q., Yang L.T., and Li Y.R., 2011, Comparison of physiological and biochemical characteristics related to cold resistance in sugarcane under field conditions, Zuowu Xuebao (Acta Agronomica Sinica), 37(3): 496-505 (张保青, 杨丽涛, 李杨瑞, 2011, 自然条件下甘蔗品种抗寒生理生化特性的比较, 作物学报, 37(3): 496-505)
Zhang C.J., Li J., Fan H.P., Wang Y.K., and Zhou D.D., 2008, A study on the hardiness of young fruits of different varieties of apricot, Xibei Linxueyuan Xuebao (Journal of Northwest Forestry University), 23(3): 29-32 (张成军, 李捷, 范惠萍, 王有科, 周丹丹, 2008, 不同杏品种幼果抗寒性研究, 西北林学院学报, 23(3): 29-32)