研究报告/Research Report
解除独行菜种子低温萌发停滞的温度响应蛋白筛选及表达分析 
2北京师范大学生命科学学院, 抗性基因资源与分子发育北京市重点实验室, 北京, 100875
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2016 年, 第 14 卷, 第 13 篇 doi: 10.5376/mpb.cn.2016.14.0013
收稿日期: 2016年04月22日 接受日期: 2016年05月25日 发表日期: 2016年05月29日
引用格式(中文):
李萍萍等, 2016, 解除独行菜种子低温萌发停滞的温度响应蛋白筛选及表达分析, 分子植物育种(online), 14(13): 1087-1094 (doi: 10.5376/mpb.cn.2016.14.0013)
引用格式(英文):
新疆北部早春短命植物独行菜(Lepidium apetalum Willd.)种子萌发对温度响应独特:低温层积可以促进种子的萌发并显著增加整齐度,但在4℃条件下存在低温萌发停滞现象,而经25℃处理45~55 min及更长时间后,种子则可以在4℃条件下萌发。这种短暂的高温诱导解除低温萌发停滞的机制尚不清楚。本研究对处于低温层积前、萌发停滞状态及25℃高温处理解除低温萌发停滞后的种子进行蛋白质差异分析,筛选出解除独行菜种子低温萌发停滞的温度响应蛋白,采用定量qRT-PCR技术对相应基因的表达进行了验证。结果表明:低温层积前、后独行菜种子之间差异蛋白不显著;25℃高温处理45 min解除低温萌发停滞的种子与前二者种子蛋白差异显著。从中筛选出37个差异蛋白,其中14个上调,23个下调。选择显著上调的6个蛋白进行LC/MS-MS鉴定,并在生物信息学分析基础上,进一步选择其中的Cdc48E、Hsp17.6和Per12进行qRT-PCR验证,结果表明:这三个蛋白的基因在解除低温萌发停滞的种子中表达量极显著增高;随25℃处理时间增加,三个基因表达均呈上升趋势,且处理在45~55 min时上调表达达到最高,之后维持这一水平表达,这与经25℃处理30~60 min能够解除低温萌发停滞进行萌发的种子比率显著增加的特性相一致。以上结果说明Cdc48E、Hsp17.6和Per12可能与解除独行菜种子低温萌发停滞有密切关系,此结果为植物温度信号响应的研究提供了新的线索。
研究背景
独行菜(Lepidium apetalum Willd.),属十字花科独行菜属一年生中生短命草本植物,其根、茎、叶有大型薄壁细胞(于喜凤和刘瑷清, 1997),且维管束有类似“花环”的结构,具有高光特效性,属C4植物(李秀明, 2009)。独行菜种子呈扁卵状,棕红色,富含脂肪,蛋白质,糖,芥子甙等,具有强心,平喘,利尿的作用。与抱茎独行菜种子类似,独行菜干种子表面覆有一层粘液质种衣,遇水后呈射线状向外发射,主要成分为多糖(谷丽丽等, 2008),能迅速地从周围土壤中吸收水分利于萌发,但对幼苗的生长基本不起作用(袁祯燕等, 2006)。生活在新疆北部荒漠的独行菜,在植物区系中为先锋植物(李岩等, 2014),具有重要的生态意义,具有典型的早春短命植物的特点,能够耐受早春低温进行萌发及幼苗生长。赵惠新等采用cDNA-AFLP技术从独行菜幼苗中筛选获得了冷上调表达的18个基因(Zhao et al., 2012),为研究独行菜幼苗耐受低温胁迫的分子基础提供了一些线索。
独行菜种子萌发对温度有特殊的响应,实验室条件下,在低温下(0℃~4℃)不萌发,在10℃~25℃能够萌发(孟君等, 2008)。而野外条件下,在新疆北部早春平均气温为2℃~3℃低温下能够萌发,表现出对低温胁迫具有较好的耐受性。独行菜为何能适应早春低温进行萌发而在实验室低温条件不能萌发,赵惠新等(2010)在研究中指出,虽然独行菜种子不能在4℃低温萌发,但种子萌发至露白前存在一个关键的生理阶段,在该阶段之前和之后均能耐受低温。当独行菜种子在早春吸水后进行萌发初期阶段,如果温度低则停滞在一个特定的萌发阶段,一旦有一定时间的温度升高,萌发的低温停滞就被打破。这与实验室适当的低温层积能显著提高独行菜种子的萌发势;4℃下,独行菜种子不能萌发,但层积10 d再经25℃处理1 h的独行菜种子就能在4℃低温下萌发的实验结果一致。另外,独行菜种子越过低温停滞后的其它不同萌发时期经冷诱导处理,对-5℃,-10℃胁迫具有较强耐受性(孟君等, 2008),为了从生理方面解析独行菜种子萌发低温耐受性的机制,杨娜等(2015)研究报道了独行菜耐受低温萌发过程中低温对超氧化物歧化酶(SOD)、过氧化氢酶(CAT)活性的影响很大。周茜等(2016)对独行菜低温萌发停滞解除前后的种子转录组进行测序,发现159个表达量存在显著差异的基因,其中表达量显著上调的基因有54个。但从蛋白水平进行独行菜种子耐受低温进行萌发的分子响应机制还没有报道。本研究就独行菜种子萌发的低温停滞及其解除的相关响应蛋白进行筛选,并选择重要的差异蛋白进行鉴定,讨论它们的功能及在萌发停滞解除过程中的基因表达模式,分析其表达量与萌发停滞的关联性,为阐明独行菜种子萌发的低温停滞机制奠定一定的基础。
1结果与分析
1.1不同萌发时期独行菜种子总蛋白含量的变化
采用TCA/丙酮沉淀法,提取低温萌发停滞解除前后独行菜种子总蛋白,对三组种子总蛋白提取及含量测定结果表明,第一组(未经低温层积)种子总蛋白含量最高,达到3.46%,可能含有大量的储存蛋白。第二组(低温层积10 d后处于萌发低温停滞期)种子总蛋白含量显著降低,为2.91%,可能是在最初的萌发期,大量的储存蛋白分解为种子萌发提供能量,或将储存蛋白转变成种子萌发需要的其他非蛋白物质。第三组(25℃处理50 min解除低温停滞)种子的蛋白含量较停滞期的种子的蛋白含量显著升高,达到3.19%,说明从种子萌发停滞到解除萌发停滞的过程中,有新的蛋白合成,可能与种子低温萌发密切相关。
1.2独行菜种子萌发低温相关响应蛋白筛选
2-DE图谱分析表明,三组种子都能检测到约600个蛋白质点。与第一、第二组进行比较,从第三组种子(解除低温萌发停滞的种子)的2D图谱中筛选显著差异的蛋白质点,共得到37个(图1),其中14个上调表达,23个下调表达。选取表达显著上调的6个蛋白点(点24、27、28、29、31、32)做LC/MS-MS质谱鉴定。鉴定结果用MASCOT在Swissprot、NCBI数据库进行比对分析,这6个蛋白分别是1个分子陪伴蛋白Cpn60,2个热激蛋白Hsp17.6I、Hsp70B,1个细胞分裂控制蛋白48家族E(Cdc48E),和2个氧化还原酶Ш家族成员过氧化物酶12(Per12)、过氧化物酶28(Per28)。分子陪伴蛋白Cpn60作为分子伴侣,参与蛋白的折叠,帮助相关蛋白发挥相应作用;当受到热激,渗透压休克或盐胁迫时,Hsp17.6I在植物种子发育和萌发过程中表达,定位于细胞质中,也起到分子伴侣的作用(Sun et al., 2001)。短暂热激后,独行菜种子中合成热激蛋白,以响应环境变化。细胞分裂控制蛋白48家族E是细胞分裂控制蛋白家族的一员,参与调控细胞分裂过程,可能参与种子萌发过程中的细胞分裂和增殖。过氧化物酶(POD)与呼吸作用,光合作用及生长素的氧化等都有关系,其生理活性随植物生长发育过程的进行而不断变化,可能参与独行菜种子低温萌发耐受。在后续的实验中,选择有分子伴侣作用的Hsp17.6、过氧化物酶Per12、细胞分裂控制蛋白Cdc48E进行表达验证。
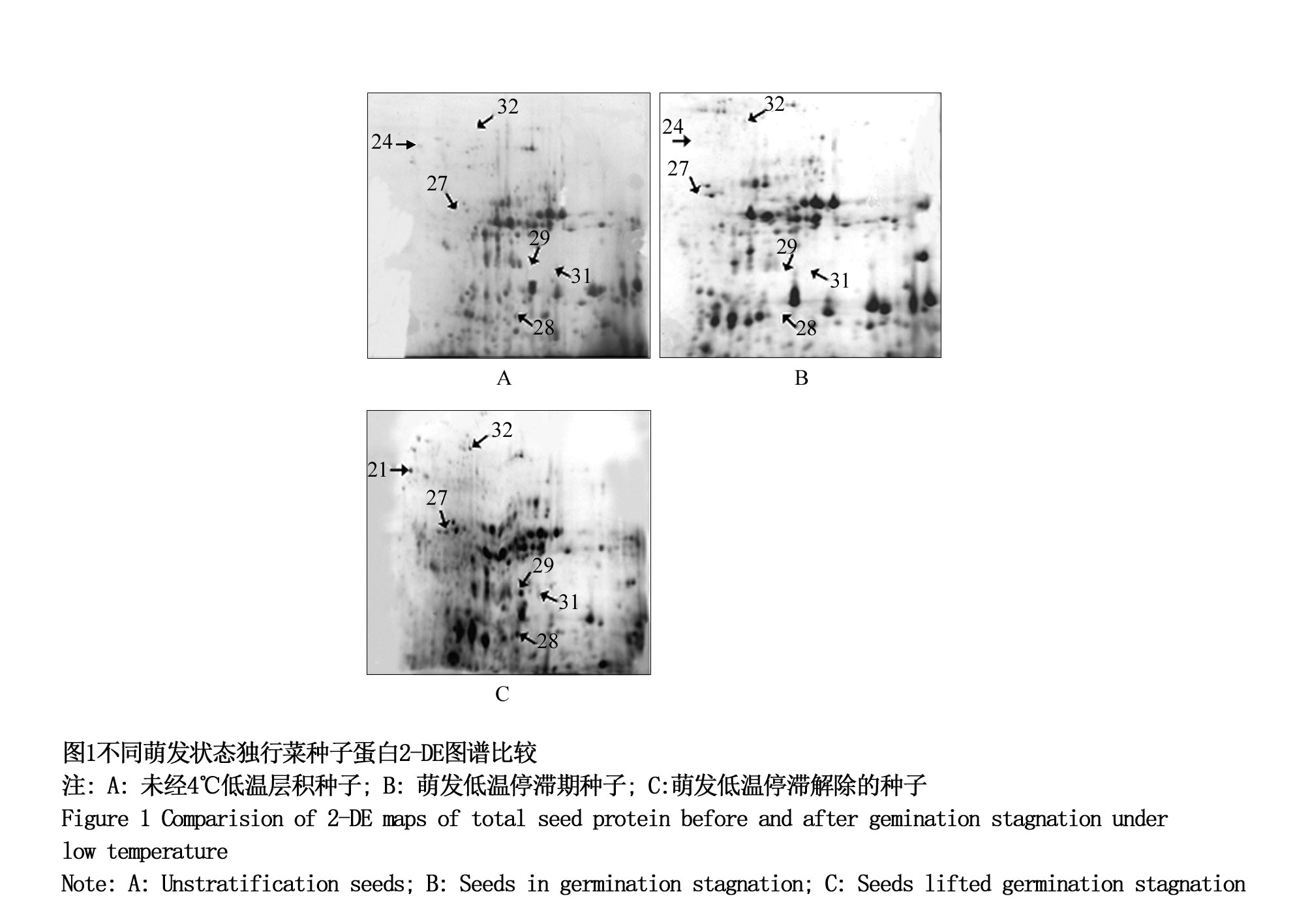 图1 不同萌发状态独行菜种子蛋白2-DE图谱比较
注: A: 未经4℃低温层积种子; B: 萌发低温停滞期种子; C:萌发低温停滞解除的种子 Figure 1 Comparision of 2-DE maps of total seed protein before and after gemination stagnation under low temperature Note: A: unstratification seeds; B: seeds in germination stagnation; C: seeds lifted germination stagnation |
1.3独行菜种子萌发低温相关响应蛋白的表达量与低温萌发关系的分析
1.3.1 CDC48E,PER12和HSP17.6基因在独行菜种子萌发低温停滞解除前后的表达量分析
为了进一步探讨筛选获得的差异表达蛋白对独行菜低温萌发停滞解除的作用,本研究选择了CDC48E,PER12和HSP17.6三个基因,比较了它们在第一、第二组、第三组种子中表达情况。结果表明,三种基因在第一组、第二组(萌发低温停滞前)的种子中表达量均极低,而在低温萌发停滞解除后CDC48E,PER12和HSP17.6基因表达量均极显著上调。说明短时间较高温度刺激可促使CDC48E,PER12和HSP17.6基因的表达,这些基因的表达可能与独行菜种子解除低温萌发停滞密切相关(图2)。
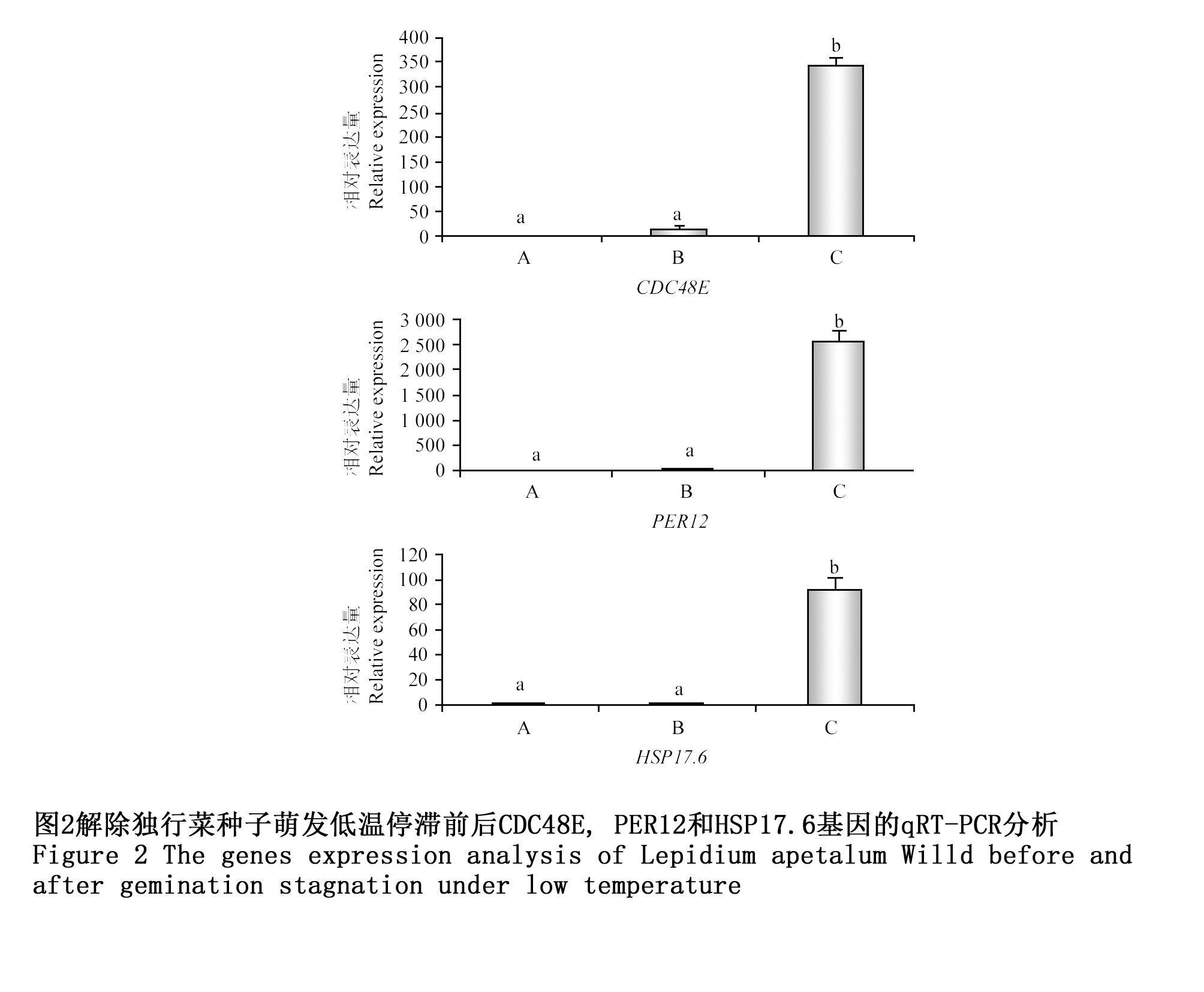 图2 解除独行菜种子萌发低温停滞前后CDC48E, PER12和HSP17.6基因的qRT-PCR分析 Figure 2 The genes expression analysis of Lepidium apetalum Willd before and after gemination stagnation under low temperature |
1.3.2 Cdc48E,Per12和Hsp17.6等低温萌发相关响应蛋白的表达量与低温萌发关系的分析
独行菜种子在4℃下层积10 d后,处于萌发停滞期。用25℃分别处理30 min,45 min,55 min,1 h,统计不同时间处理对独行菜种子耐受低温萌发率的影响。经25℃,30 min,45 min,55 min,1 h诱导处理处于萌发停滞期的独行菜种子后,在4℃条件下萌发率分别达到0%、40%,83.33%,88.33%,96.11%;也就是经30 min的25℃处理低温停滞期的种子后,40%的种子解除萌发停滞,耐受低温继续进行萌发;经45 min的25℃处理,83.33%的种子解除萌发停滞;经1 h的25℃处理,种子耐受低温萌发率达到正常温度萌发率,也就是几乎所有的种子能够解除萌发停滞(图3)。
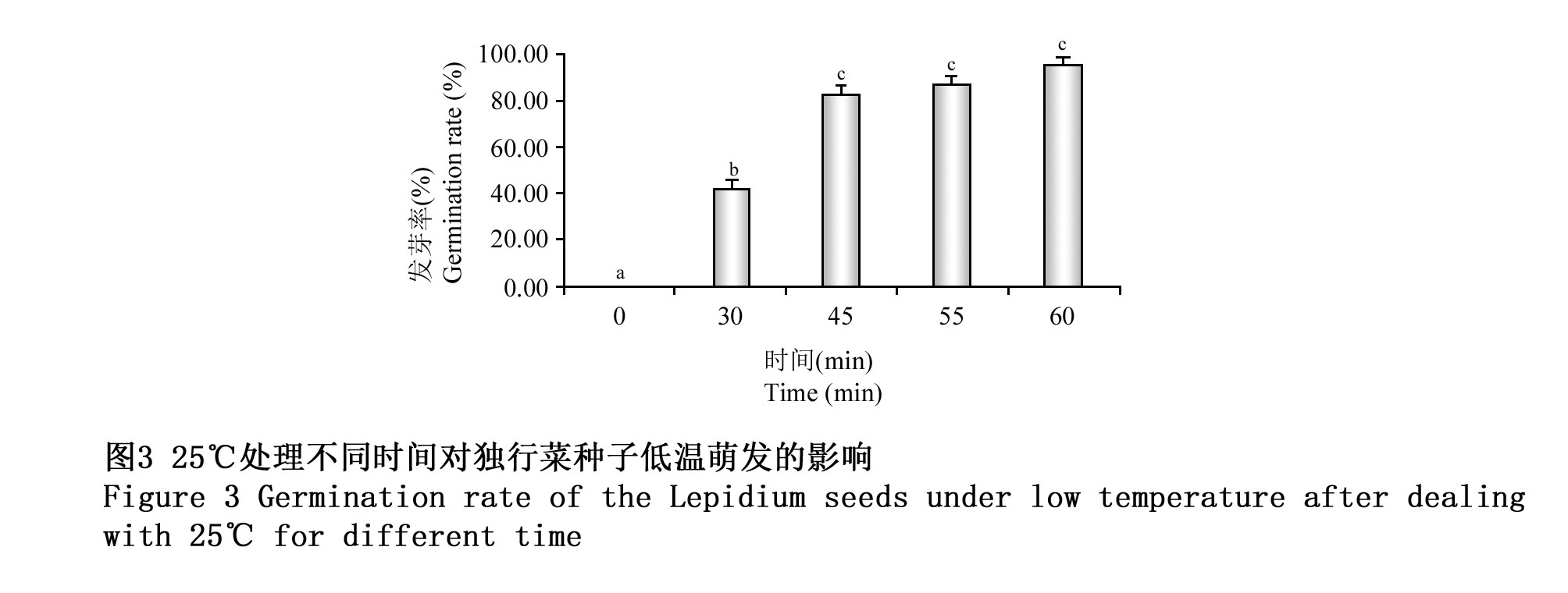 图3 25℃处理不同时间对独行菜种子低温萌发的影响 Figure 3 Germination rate of the Lepidium seeds under low temperature after dealing with 25℃ for different time |
为进一步探讨筛选获得的相关响应蛋白表达与独行菜低温萌发间的关系,对CDC48E,PER12和HSP17.6三个基因在经不同时间25℃诱导处理处于萌发低温停滞期的种子中的表达情况进行分析。25℃处理30 min后,与处于低温停滞期的独行菜种子相比较,CDC48E,PER12和HSP17.6基因的表达量均极显著提高(p<0.01),且在1 h内随处理时间的增加均呈上升趋势(图4)。三个基因的表达特性也有一定的差异,其中CDC48E、PER12基因经25℃处理不同时间均有极显著上调;而HSP17.6基因表达量在25℃处理50~60 min时极显著上调(p<0.01),而在25℃处理45 min以下显著上调表达(p<0.05)。这与经25℃处理超过45 min后,80%以上的种子都能解除萌发停滞期,耐受低温继续进行萌发的变化趋势一致。这也说明筛选获得的25℃诱导上调表达的蛋白对独行菜种子低温萌发停滞解除关系密切。
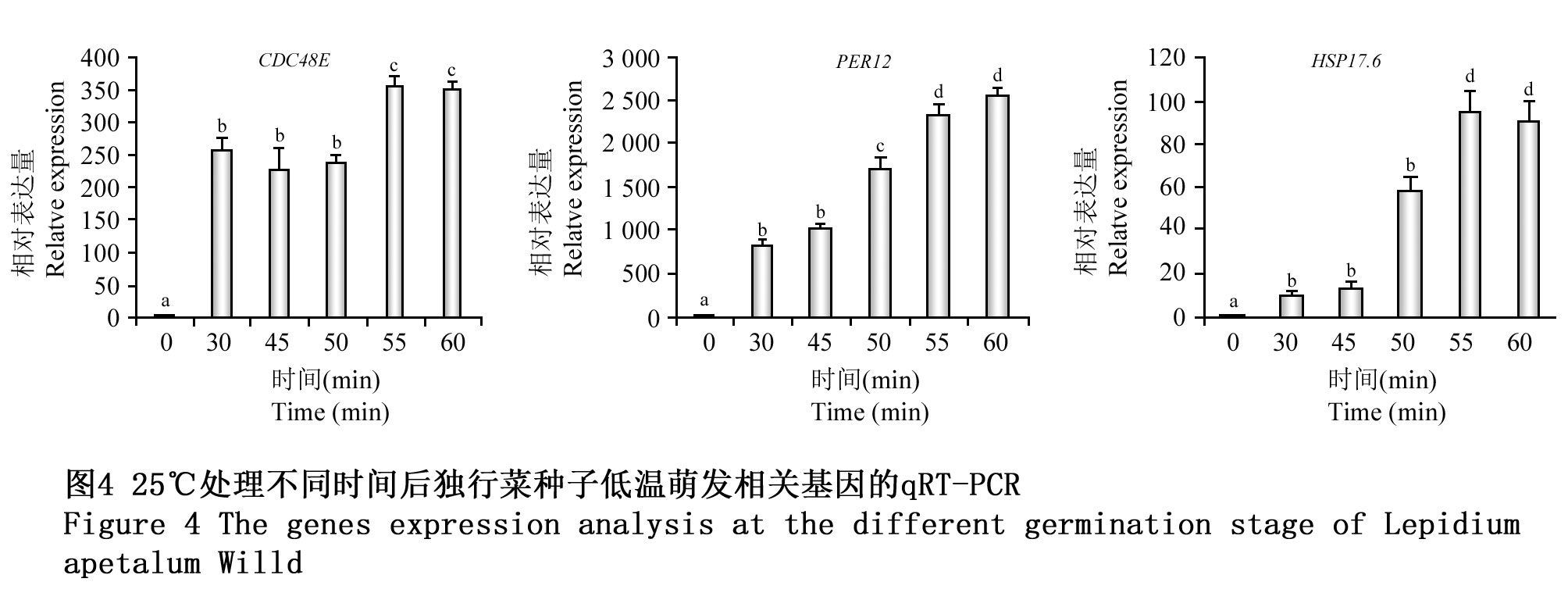 图4 25℃处理不同时间后独行菜种子低温萌发相关基因的qRT-PCR Figure 4 The genes expression analysis at the different germination stage of Lepidium apetalum Willd. |
2讨论
种子萌发是一个非常复杂的过程,涉及到许多与生长发育相关的蛋白的调控表达。本研究以生活在新疆北部的独行菜种子这一特殊生态型为研究材料,通过比较其低温萌发停滞解除前后蛋白表达的差异,筛选出停滞解除表达量显著上调的蛋白并进行质谱鉴定分析,并对其中部分蛋白做了相应的基因表达分析。
热激蛋白是一类在系统发育过程中高度保守的分子伴侣,按相对分子量(KD)大小可以分为Hsp100s,Hsp90s,Hsp70s,Hsp60s和sHsps。前四种Hsp被视为HWM Hsps (high molecular weight Hsps),它们主要参与一些蛋白的折叠和装配。Hsp70B定位在细胞质膜,叶绿体和细胞质中,与其它分子伴侣协同作用,参与胞质溶胶和细胞器中新生蛋白的富集和折叠是热激诱导合成的,能减轻植物在冷环境中的膜伤害,增强组织的耐冷性,与植物获得耐受性关系密切(Lin et al., 2001),黄上志等发现萌发的水稻种子经42℃热激处理后其幼苗的耐冷性明显增强(黄上志等, 2004)。细胞质中分子陪伴蛋白Cpn60-α1亚基可以结合Rubisco大亚基(陈根云, 2000),帮助蛋白正确折叠,免受其他蛋白的干扰。Hsp17.6I在独行菜种子解除低温萌发停滞的过程中,表达量增加,提高了其耐冷性。
过氧化物酶(POD)是植物细胞内的保护酶,不仅与植物抵御低温等不良环境有关,还在一定温度范围内,随温度升高,活性增加(刘应迪等, 2001)。Per12和Per28都是典型的氧化还原酶Ш家族成员(Tognolli et al., 2002),属于跨膜蛋白,主要存在于液泡中,在种子发育早期受环境诱导表达,它有许多酶辅助因子结合位点,如血红素b和Ca2+结合位点(Theologis et al., 2000; Dunand et al., 2002)。当植物组织受到外界环境压力如创伤,病原体侵害和氧化应激时,可以依赖各同工酶亚型清除H2O2,有毒还原剂,合成和降解木质素,木栓,分解代谢生长素等(Schenk et al., 2000)。短时间较高温度刺激低温层积的独行菜种子,种子内过氧化物酶含量增加,利于H2O2的清除,提高其耐受低温的能力。然而,在种子萌发初期,通过抑制某些过氧化物酶基因的表达量,如抗坏血酸过氧化物酶(APX)(Sung et al., 2005),可以使种子内的H2O2得到积累,从而刺激种子的萌发,提高萌发率。故而,独行菜种子低温萌发停滞解除过程中,过氧化物的增加虽保护了独行菜免受低温伤害,但不是促进其萌发的关键基因。
细胞分裂控制蛋白E(Cdc48E)属于AAAA- TPase家族,亚细胞定位主要在细胞核中,随着细胞分裂过程中新细胞壁的形成定位于成膜体中,通过与相同的细胞器发生同型膜融合从而在细胞分裂和生长过程中起作用(Tabata et al., 2000; Seki et al., 2002)。Cdc48E通常包含809个氨基酸,在拟南芥的芽等生长旺盛的细胞中高度表达,参与新细胞壁的形成和核膜的解体与重建(Yamada et al., 2003; Saracco et al., 2009)。25℃处理低温层积10 d的独行菜种子1 h后,Cdc48E蛋白含量显著增加,促进细胞周期进行,有利于胚细胞进一步的生长和发育。
本研究分别从蛋白水平和转录水平对独行菜种子低温萌发不同时期的基因表达情况进行了分析和讨论。结果表明:细胞分裂控制蛋白E基因(CDC48E),过氧化物酶12基因(PER12)和小分子量热激蛋白基因(HSP17.6)在低温层积的独行菜种子经25℃处理一段时间后,显著上调表达,这与蛋白表达结果一致。25℃处理时间的长短影响萌发停滞期的独行菜种子的萌发率,且萌发率随处理时间的增长明显提高。这一表型变化结果也佐证了25℃短时间处理低温层积的独行菜种子,会影响某些基因的表达,而这些基因的表达产物对独行菜耐受低温萌发起到有利作用。低温是影响作物产量的关键逆境因子之一,研究影响独行菜种子低温萌发的关键因子,可以帮助了解植物低温萌发的分子机制,最终为改良作物的遗传形状提供依据。
3材料与方法
3.1材料及处理
独行菜种子采集于新疆乌鲁木齐市鲤鱼山,选择成熟饱满的种子,室温干燥保存。
通过对不同温度打破低温抑制的效果进行比较,确定25℃的效果最好(赵惠新等, 2010)。种子处理分为三组:第一组是低温层积前的种子,即直接用无菌水湿润的种子;第二组是低温萌发停滞状态的种子,即湿润后的种子置于4℃层积10 d;第三组是解除低温萌发停滞的种子,即层积后的种子置于25℃处理50 min。种子样品液氮条件下研磨成粉末,置于-80℃冰箱备用。
3.2蛋白质样品的制备及蛋白含量测定
采用TCA/丙酮沉淀法(Agnieszka and Juan, 2013; Vasconcelos et al., 2005; Wu et al., 2014)分别提取一、二、三组种子的总蛋白。采用Bradford法(1976)测定总蛋白的浓度,计算各样品蛋白含量。
3.3独行菜种子萌发低温相关响应蛋白的筛选
采用双向电泳技术分别对三组蛋白质进行分离。其中等电聚焦选用17 cm线性IPG预制胶条(pH3-10),上样量按照种子质量统一进行定量上样,具体方法参照野生小麦(Gharechahi et al., 2014)和春大豆(Tian et al., 2015 )蛋白的双向电泳过程,略作改动。与第一、第二组进行比较,从第三组种子(解除低温萌发停滞的种子)样品中筛选差异表达的蛋白。运用PDQuest8.0软件分析2-DE图谱,筛选表达量变化超过3倍的蛋白质点,并采用LC/MS-MS进行质谱鉴定。
3.4独行菜种子萌发低温相关响应蛋白的表达量与低温萌发关系的分析
比较筛选获得的几种与低温萌发响应相关蛋白在第一、第二组、第三组种子中表达情况;将处于低温萌发停滞期的种子分成5份,分别用25℃诱导处理0、30 min、45 min、50 min、55 min、1 h,用于差异蛋白基因表达的RT-PCR分析验证。并对部分种子继续放置于4℃进行萌发实验,15 d后统计萌发率。综合实验结果对独行菜种子萌发低温相关响应蛋白的表达量与低温萌发关系进行分析。
筛选获得的蛋白表达情况采用荧光定量RT-PCR进行检测,具体操作如下:采用Trizol法(TIANGEN)提取独行菜种子总RNA。参照Revert Aid First Strand cDNA Synthesis Kit (Thermo)操作说明书对上述RNA样本进行第一链cDNA的合成。根据MASCOT搜索结果,利用Primer5.0根据目标蛋白相应核苷酸序列设计引物,引物序列及退火温度和扩增产物大小(表1)。以ACTIN为内参基因,利用UltraSYBR Mixture(With ROX II)(康为世纪)对目的基因进行表达分析。每个样品做三次生物学重复。
.png) 表1 关键蛋白qRT-PCR分析的引物序列及预期产物大小 Table 1 Sequences of the primers used in qRT-PCR analysis for key protein and the product size |
作者贡献
李萍萍是实验研究、数据分析的主要执行人,并完成论文初稿的写作;赵惠新、赵和平指导论文写作和修改;曾卫军、赵和平进行中英文摘要的修改;曾卫军、赵和平、李艳红、葛风伟、祝长青指导实验设计和数据分析;卢函,周茜和赵君洁参与实验实施,实验结果分析,图表处理;赵惠新是实验设计者、项目的构思者及负责人。全体作者都阅读并同意最终的文本。
致谢
本研究由国家自然科学基金(31460041)、新疆维吾尔自治区教育厅重点实验室项目(XJTSWZ- 2015-01)、抗性基因资源与分子发育北京市重点实验室开放课题(2015GD03)和新疆师范大学博士启动基金项目(XJNUBS1416)共同资助。
Agnieszka Z., and Juan D.R., 2013, A protocol for protein extraction from lipid-rich plant tissues suitable for electrophoresis, Plant Proteomics, 1072: 85-91
Bradford M.M., 1976, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding, Analytical Biochemistry, 72: 248-254
http://dx.doi.org/10.1016/0003-2697(76)90527-3
Chen G.Y., Xiao Y.Z., and Li L.R., 2000, Rubisco assembly in higher plant and characterization of its intermediates, Zhiwu Shengli Xuebao (Acta Phytophysiological Sinica), 26(1): 16-22 (陈根云, 肖元珍, 李立人, 2000, 高等植物Rubisco的组装及其中间产物的鉴定, 植物生理学报, 26(1): 16-22)
Dunand C., Tognolli M., and Overney S., 2002, Identification and characterization of Ca(2+)-pectate binding peroxidases in Arabidopsis thaliana, Plant Physiol, 159(11): 1165-1171
http://dx.doi.org/10.1078/0176-1617-00768
Gharechahi J., Alizadeh H., Naghavi M.R., and Sharifi G., 2014, A proteomic analysis to identify cold acclimation associated proteins in wild wheat(Triticum urartu L.), Mol Biol Rep, 41(6): 3897-3905
http://dx.doi.org/10.1007/s11033-014-3257-8
PMid:24535272
Gu L.L., Liu L.H., You T.Y., Lan H.Y., and Zhang F.C., 2008, Characterization of the seed coat mucilage properties of ephemeral plant Lepidium perfoliatum L. in Xinjiang, Xibei Zhiwu Xuebao (Acta Botanica Boreali-Occidentalia Sinica), 28(12): 245-246 (谷丽丽, 刘立鸿, 油天钰, 兰海燕, 张富春, 2008, 新疆短命植物抱茎独行菜种子粘液质特性的研究(英文), 西北植物学报, 28(12): 245-246)
Huang S.Z., Huang X.F., Lin X.D., Zhang Y.S., Liu J., and Fu J.R., 2004, Induction of chilling tolerance and heat shock protein synthesis in rice seedlings by heat shock, Zhiwu Shengli Yu Fenzi Shengwuxue Xuebao (Journal of Plant Physiology and Molecular Biology), 30(2): 189-194 (黄上志, 黄祥富, 林晓东, 张以顺, 刘军, 傅家瑞, 2004, 热激对水稻幼苗耐冷性及热激蛋白合成的诱导, 植物生理与分子生物学学报, 30(2): 189-194)
Li X.M., Liu P., Gu L.L., Yao S.X., You T.Y., Lan H.Y., and Zhang F.C., 2009, Studies on the C4-related structures of two ephemerals-eremopyrum orientale(L.) and Lepidium apetalum Willd. in Xinjiang, Xinjiang Nongye Kexue (Xinjiang Agricultural Sciences), 46(1): 28-33 (李秀明, 刘彭, 谷丽丽, 姚世响, 油天钰, 兰海燕, 张富春, 2009, 新疆短命植物东方旱麦草和独行菜C4相关结构的初步研究, 新疆农业科学, 46(1): 28-33)
Li Y., Feng Y., Liu B., Lv G.H., and Wang X.Y., 2014, Phylogenetic relationships and divergence time of Brassicaceae ephemeral plants in Xinjiang, Ganhanqu Yanjiu (Arid Zone Research), 31(6): 1100-1108 (李岩, 冯缨, 刘斌, 吕光辉, 王习勇, 2014, 新疆十字花科短命植物系统发育关系及分化时间, 干旱区研究, 31(6): 1100-1108)
Lin B.L., Wang J.S., and Liu H.C., 2001, Genomic analysis of the Hsp70 superfamily in Arabidopsis thaliana, Cell Stress Chaperones, 6(3): 201-208
http://dx.doi.org/10.1379/1466-1268(2001)006<0201:GAOTHS>2.0.CO;2
Liu Y.D., Cao T., Xiang F., and Peng C.H., 2001, Effect of high temperature stress on the activity of peroxidase of two species of mosses, Guangxi Zhiwu (Guihaia), 21(3): 255-258 (刘应迪, 曹同, 向芬, 彭春晖, 2001, 高温胁迫下两种藓类植物过氧化物酶活性的变化, 广西植物, 21(3): 255-258)
Meng J., Li Q., and Li G., 2008, Physiological characteristic of seed germination of two species of Lepidium L, Shengwu Jishu (Biotechnology), 18(2): 32-35 (孟君, 李群, 李冠, 2008, 两种独行菜种子萌发生理特性研究, 生物技术, 18(2): 32-35)
Saracco S.A., Hansson M., and Scalf M., 2009, Tandem affinity purification and mass spectrometric analysis of ubiquitylated proteins in Arabidopsis, Plant J, 59(2): 344-358
http://dx.doi.org/10.1111/j.1365-313X.2009.03862.x
PMid:19292762 PMCid:PMC3639010
Schenk P.M., Kazan K., Wilson I., Jonathan P.A., Todd R., Shauna C.S., and John M.M., 2000, Coordinated plant defense responses in Arabidopsis revealed by microarray analysis, Proc. Natl. Acad. Sci. U.S.A., 97(21): 11655-11660
http://dx.doi.org/10.1073/pnas.97.21.11655
PMid:11027363 PMCid:PMC17256
Seki M., Narusaka M., Kamiya A., Ishida J., Satou M., Sakurai T., Nakajima M., Enju A., Akiyama K., Oono Y., Muramatsu M., Hayashizaki Y., Kawai J., Carninci P., Itoh M., Ishii Y., Arakawa T., Shibata K., Shinagawa A., and Shinozaki K., 2002, Functional annotation of a full-length Arabidopsis cDNA collection, Science, 296(5565): 141-145,
http://dx.doi.org/10.1126/science.1071006
PMid:11910074
Sun W., Bernard C., van de Cotte B., Montagu M.V., and Verbruggen N., 2001, At-HSP17.6A, encoding a small heat-shock protein in Arabidopsis, can enhance osmotolerance upon overexpression, Plant J, 27(5): 407-415
http://dx.doi.org/10.1046/j.1365-313X.2001.01107.x
PMid:11576425
Sung C.M., Soon I.K., and Chung S.A., 2005, Changes in expression of the cytosolic ascorbate peroxidase gene, Ca-cAPX1, during germination and development of Hot Pepper seedlings, Journal of Plant Biology, 48(3): 276-283.
http://dx.doi.org/10.1007/BF03030523
Tabata S., Kaneko T., Nakamura Y., et al., 2000, Sequence and analysis of chromosome 5 of the plant Arabidopsis thaliana, Nature, 408(6814): 823-826
http://dx.doi.org/10.1038/35048507
PMid:11130714
Theologis A., Ecker J.R., Palm C.J., et al., 2000, Sequence and analysis of chromosome 1 of the plant Arabidopsis thaliana, Nature, 408(6814): 816-820
http://dx.doi.org/10.1038/35048500
PMid:11130712
Tian X., Liu Y., Huang Z.G., Duan H.P., Tong J.H., He X.L., Gu W.H., Ma H., and Xiao L.T., 2015, Comparative proteomic analysis of seedling leaves of cold-tolerant and -sensitive spring soybean cultivars, Mol Biol Rep, 42(3): 581-601
http://dx.doi.org/10.1007/s11033-014-3803-4
PMid:25359310
Tognolli M., Penel C., Greppin H., and Simon P., 2002, Analysis and expression of the class III peroxidase large gene family in Arabidopsis thaliana, Gene, 288(s 1-2): 129-138
Vasconcelos É.A.R., Nogueira F.C.S., and Abreu E.F.M., 2005, Protein extraction from cowpea tissues for 2-D gel electrophoresis and MS analysis, Chromatographia, 62(7-8): 447-450
http://dx.doi.org/10.1365/s10337-005-0637-1
Wu X.L., Gong F.P., and Wang W., 2014, Protein extraction from plant tissues for 2DE and its application in proteomic analysis, Proteomics, 14(6): 645-658
http://dx.doi.org/10.1002/pmic.201300239
PMid:24395710
Yamada K., Lim J., Dale J.M., Chen H.M., Shinn P., Palm C.J., et al., 2003, Empirical analysis of transcriptional activity in the Arabidopsis genome, Science, 302(5646): 842-846
http://dx.doi.org/10.1126/science.1088305
PMid:14593172
Yang N., Zhao H. P., Ge F.W., Li Y.H., Zeng W.J., and Zhao H.X., 2015, Physiological response of two Lepidium species to low temperature stress during seed germination, Ganhanqu Yanjiu (Arid Zone Research), 32(4): 760-765 (杨娜, 赵和平, 葛风伟, 李艳红, 曾卫军, 赵惠新, 2015, 2种独行菜萌发对低温胁迫的生理响应, 干旱区研究, 32(4): 760-765)
Yu X.F., and Liu A.Q., 1997, A anatomical study on the ephemeral plant-Lepidium Apetalum Willd. in XinJiang, Xinjiang Shifan Daxue Xuebao (Journal of Xinjiang Normal University(Natural Science Edition)), 02: 34-38(于喜凤, 刘瑷清, 1997, 新疆"短命植物"独行菜营养器官的解剖学科研究, 新疆师范大学学报(自然科学版), 02: 34-38)
Yuan Z.Y., Duo L.K.•M.M.T.Y.S.F., and Huang P.Y., 2006, The relationship between water and the seed coat of Lepidium Apetalum- an ephemeral, Zhongzi (Seed), 25(9): 1-3 (袁祯燕, 多力坤•买买提玉素甫, 黄培佑, 2006, 早春短命植物独行菜天然种衣与水分的关系, 种子, 25(9): 1-3)
Zhao H.X., Li Q., LiG., and Du Y., 2012, cDNA-AFLP Analysis reveals differential gene expression in response to cold stress in Lepidium apetalum during seedling emergence, Biologia Plantarum, 56(1): 64-70
http://dx.doi.org/10.1007/s10535-012-0017-2
Zhao H.X., Li Q., Zhou J., and Li G., 2010, The characteristics of low temperature tolerance during seed gemination of the ephemeral plant Lepidium apetalum (Cruciferae), Yunnan Zhiwu Yanjiu (Acta Botanica Yunnanica), 32(5): 448-454 (赵惠新, 李群, 周晶, 李冠, 2010, 短命植物独行菜种子萌发过程对低温的耐受特性, 云南植物研究, 32(5): 448-454)
Zhou X., Li P. P., Zeng W. J., Li Y. H., Ge F. W., Yang N., Zhao H. X., 2016, De novo characterization of the seed transcriptome of Lepidium apetalum Willd., Zhongguo Shengwu Gongcheng Zazhi (China Biotechnology), 01: 1-11 (周茜, 李萍萍, 曾卫军, 李艳红, 葛凤伟, 杨娜, 赵惠新, 2016, 独行菜种子转录组的高通量测序及分析, 中国生物工程杂志, 01: 1-11)