几种果树及拟南芥开花基因LFY密码子偏好性分析 
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2011 年, 第 9 卷, 第 94 篇 doi: 10.5376/mpb.cn.2011.09.0094
收稿日期: 2011年06月20日 接受日期: 2011年07月21日 发表日期: 2011年07月25日
引用格式(中文):
张颖等, 2011, 几种果树及拟南芥开花基因LFY密码子偏好性分析, 分子植物育种(online) Vol.9 No.94 pp.1673-1679 (doi: 10.5376/mpb.cn.2011. 09.0094)
引用格式(英文):
Zhang et al., 2011, Characterization of Codon Usage of LFY Genes in Several Fruit Trees, Fenzi Zhiwu Yuzhong (online) (Molecular Plant Breeding) Vol.9 No.94 pp. 1673-1679 (doi: 10.5376/mpb.cn.2011.09.0094)
由于不同物种在密码子的使用上存在不同的偏性,而密码子偏性与核酸遗传信息和生物功能载体蛋白质相关联,具有重要的生物学意义。本研究运用相关生物学软件和数理统计学软件对甜橙、桃、葡萄、芒果、甜杏、苹果、砂梨LFY (terminal flower)基因的密码子用法进行了分析,并与模式植物拟南芥的LFY基因进行了比较。结果表明,几种果树间的LFY基因密码子偏性差异不大,都偏好使用CG结尾的密码子,而拟南芥密码子第三位碱基的分布较均匀。不同物种的LFY基因在密码子上的差异与其亲缘关系呈正相关,表现为亲缘关系越近的物种其基因密码子的用法越相似。基于密码子用法的偏好研究结果可以作为物种进化关系和分子机理研究的重要补充。
生物在遗传过程中通过“中心法则”传递遗传信息,DNA的遗传信息传递给蛋白质,从而实现生物体的整个生命过程,这一过程是通过三个碱基的密码子对应一个氨基酸的方式来完成遗传信息的传递。每种氨基酸至少对应一个密码子,编码同一种氨基酸的密码子称为同义密码子。生物体内普遍存在同义密码子非均衡使用的现象,即某一物种或某一基因通常倾向于使用一种或几种特定的同义密码子,这些密码子被称为最优密码子,这一现象被称为密码子偏性,在进行基因的异源表达时,密码子偏性就显的非常重要。在分子进化、翻译调控方面,通过对果蝇、禾本科类等物种的研究表明,不同物种基因中同义密码子使用偏好性的形成与转录相关突变和翻译水平上的选择有关(Wong et al., 2002; Powell et al., 2003)。
早期关于密码子偏性的理论(如: 密码子的选择模式在进化过程中是保守的; 密码子使用的偏爱与相应的同工tRNA浓度相关, 从而有利于翻译的效率和准确性),其数据支持多来源于单细胞的大肠杆菌和酵母(Ikemura, 1985; 吴宪明等, 2007),随着新的物种被测序,大量的核酸序列和蛋白质序列被发现,研究发现多细胞生物高度的细胞分化和复杂的基因表达调控系统使密码子在选择模式上与单细胞生物存在很大差异,使密码子偏性研究进入新的发展阶段。Donal A Hickey对水稻全基因组密码子进行了分析,结果表明基因的多样性与其含有的核苷酸类别相关,特别与GC含量相关,同时密码子的偏好性与基因长度成负相关(Wang and Hickey, 2007)。
LFY (terminal flower)基因是PEBPs (phosphate dylethanolamine- binding proteins, 磷脂酰乙醇胺结合蛋白)基因家族中的一员,最初是在拟南芥中发现LFY基因在花芽形成过程及花序建成方面起到重要调控作用(Shannon and Meeks-Wagner, 1993; Bradley et al., 1997; Ratcliffe et al., 1998),随后在许多开花植物中发现LFY基因,在主要的几种果树中也陆续发现了LFY基因的同源序列,如葡萄(Carmona et al., 2002)、苹果、砂梨、枇杷(Esumi et al., 2005)和芒果(NCBI)等。果树的花芽分化是开花结果的基础,开花基因是花芽分化的内在因素,开展花芽分化相关基因的研究,阐明其表达机理,对利用分子生物学技术提高果树成花数量和质量具有重要意义。本论文从生物信息学角度分析果树LFY基因的密码子偏性,为生物进化和异源基因表达提供理论基础。
1结果与分析
1.1 LFY基因有效密码子及密码子组成
为了研究不同果树LFY基因的密码子使用方式,把来自8种果树及拟南芥的LFY基因片段的长度、ENC值、GC含量和59个密码子的RSCU值分别列表(表1; 表2)。由表2可知,不同果树LFY基因的的ENC值为52.274~56.228,都在50以上,反映了已知几种果树的LFY基因均一性较好,且与拟南芥的ENC值56.404相近。同时GC3s值进一步表明了同义密码子使用方式的均一性,表2中GC3s值范围为0.526~0.645,标准偏差为0.038,同样说明这几种果树LFY基因均一性较好,偏好性不强,稍高于拟南芥的0.497。
 表1 开花相关基因LFY编码的GC含量及有效密码子数(ENC) |
.png) 表2 开花相关基因LFY编码氨基酸的同义密码子中相对使用频率(RSCU) |
基因的表达水平越高,要求的翻译效率就越高,同时密码子的偏好性就越高,相反基因的表达水平越低,要求的翻译效率越低,密码子的偏好性也就随之降低,所以密码子偏好性的高低可以从一定程度上反映该基因的表达水平。CAI的值越低,基因越倾向于低水平表达,当CAI的值低于0.25时,mRNA水平与蛋白质水平的相关性很低(大多数基因如此),CAI大于0.5时,基因为高表达,mRNA水平与蛋白质水平的相关性较高(Carbone et al., 2003)。CAI值从0.195到0.225,平均值为0.212,其中桃和甜杏的CAI值为0.195和0.197,最高的甜橙也只有0.225,所以该基因在果树中为低水平表达,拟南芥的CAI值为0.247,同样为低水平表达(表1)。
1.2 LFY同义密码子的使用频率
利用Codon W软件对LFY基因密码子使用的具体情况进行了分析(表2),其中,甜橙RSCU值大于1的密码子有26个,桃有25个,葡萄有25个,芒果有27个,枇杷有29个,甜杏有26个,苹果28个,砂梨有27个,拟南芥有25个(7个以U结尾, 12个以A结尾)。RSCU值大于1的密码子为基因的偏爱密码子,果树中LFY基因RSCU值大于1的密码子多以C、G结尾,拟南芥中LFY基因多偏好以AU结尾的密码子(表3)。
 表3 欧氏距离系数检测开花相关基因LFY间相对密码子相对使用频率 |
1.3 LFY基因密码子使用频率的差异
根据表3中各物种密码子的相对使用频率(RSCU),利用SPSS分析软件中的欧氏距离算法,对几种果树及拟南芥的LFY基因的密码子偏好进行了计算(表4)和聚类分析(图1)。各物种间的距离系数能够体现它们在密码子使用上的相似度,由表4可知,桃和甜杏的距离最小,为 1.240;苹果和砂梨的距离也较小,为1.313;而拟南芥与所有果树的距离都较大,这一结果与图1的结果相符。基于LFY蛋白质序列构建的系统发育树的分支结果与图1的结果相一致,说明密码子的偏性差异与物种的亲缘关系存在一定的相关性,亲缘关系越近,密码子偏性差异越小。
 Table 4 The accession number of LFY genes of fruit tree and Arabidopsis thaliana |
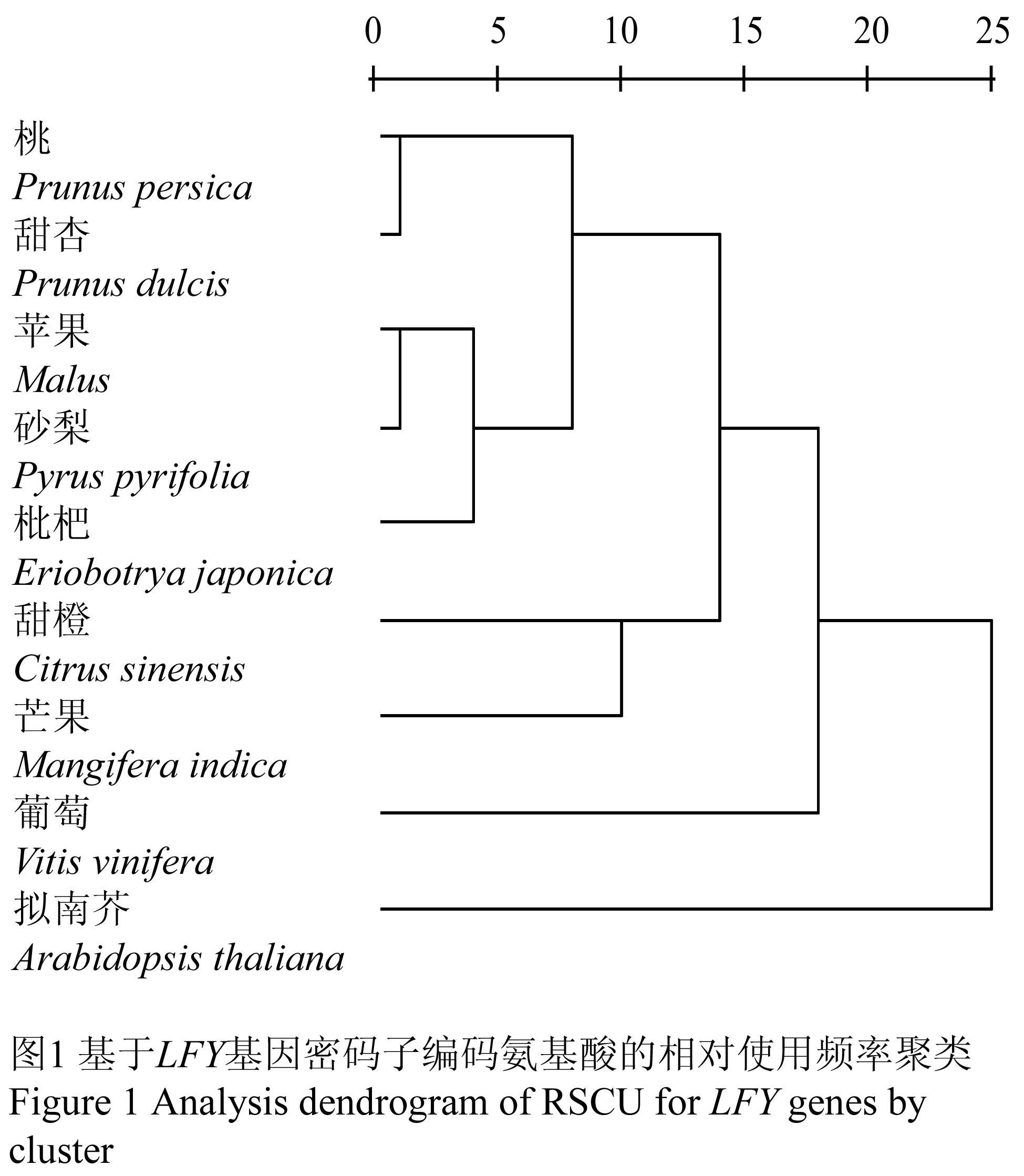 图1 基于LFY基因密码子编码氨基酸的相对使用频率聚类 Figure 1 Analysis dendrogram of RSCU for LFY genes by cluster |
2讨论
密码子使用偏性产生的生物学基础目前还不太清楚,关于偏性与翻译效率也有一些不同的看法,Kanaya等(2001)通过5个真核生物(酵母, 线虫, 果蝇, 非洲爪蟾和人)的密码子使用模式进行对比发现,密码子偏性有可能和基因组中的isochore结构和含GC二核苷酸的密码子使用有关;在植物中密码子的偏性分析也得到了类似的结论(Murray et al., 1989);zhang和Li (2004)认为基因表达的水平是决定密码子使用偏好的主要因素。
在分析GC含量的研究中,以60%为一个分界点,高于60%的为高GC含量,低于60%为低GC含量(Wang and Hickey, 2007)。本文对几个果树开花基因LFY的密码子进行偏性分析,发现几种果树的LFY基因的GC含量均低于60%,和拟南芥一起属于低GC含量基因(Wang and Hickey, 2007)。LFY基因中密码子RSCU值大于1,同时偏好一致的氨基酸所对应的密码子有如下几种:编码氨基酸Phe的密码子UUC、编码氨基酸Leu的密码子 UUG/CUU、编码氨基酸Tyr的密码子UAC、编码氨基酸Gln的密码子CAG、编码氨基酸Arg密码子AGG、编码氨基酸Thr的密码子ACG、编码Lys的密码子AAG、编码氨基酸Val的密码子GUG、编码氨基酸Glu的密码子GAG、编码氨基酸Gly的密码子GGG (表2),与拟南芥的密码子偏好特征不同(拟南芥中LFY基因多偏好以AU结尾的密码子)。
基因完整编码序列中的密码子用法在生物界广泛存在,是分子进化中的一种重要现象。密码子是携带DNA遗传信息的最小单位,从密码子偏性的特征角度可以对基因甚至物种的进化关系进行推测,不同物种或同一物种的不同基因对密码子的使用模式即密码子偏性有所不同,这种差异与物种亲缘关系存在密切关系,亲缘关系越远,密码子偏性差异大,亲缘关系近则差异小(吴宪明等, 2007; 赵薇等, 2010)。在本文中几种果树密码子偏性之间的大小(表3; 图1)与其进化关系(图2)远近相符,基于密码子使用模式差异进行的聚类分析,显示同一基因不同物种之间的关系与通过蛋白质氨基酸序列构建的系统进化树显示的各物种间的进化距离一致,说明在基因功能和类型一致的条件下,进化上亲缘关系较近的物种,密码子的使用模式相似。目前,基于分子序列的系统发育分析中,密码子使用信息的分析结果可从另外的角度为系统发育研究提供更多的参考信息,作为各种系统发育分析方法的重要补充。
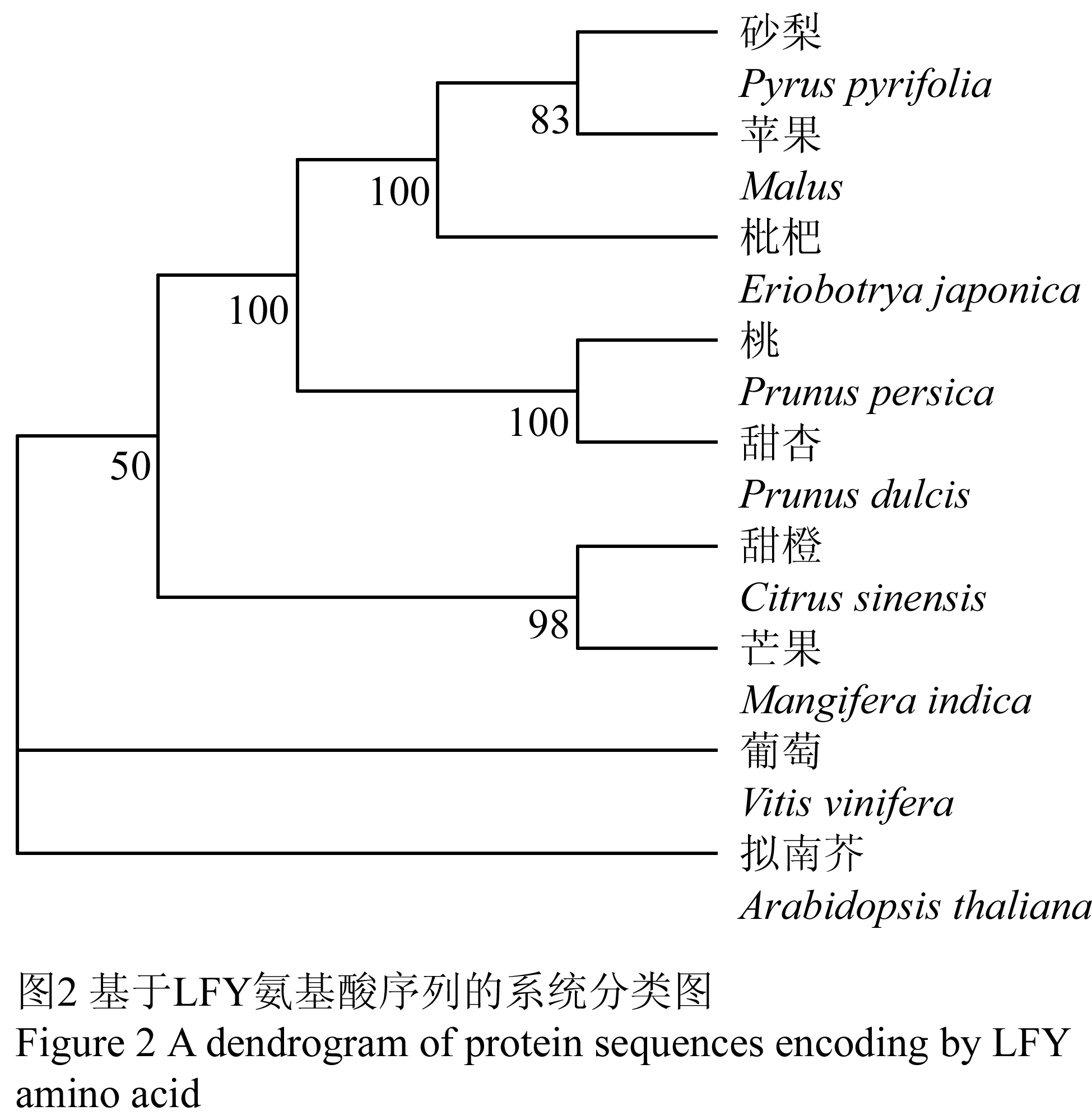 图2 基于LFY 氨基酸序列的系统分类图 Figure 2 A dendrogram of protein sequences encoding by LFY amino acid |
3材料与方法
3.1序列来源
本研究所采用的8种果树及拟南芥的LFY基因序列均来源于GenBank (表4)。
3.2使用主要软件和程序
密码子分析软件Codon W (http://www.molbio1.ox.ac.uk)和序列比对软件Invitrogen Vector NTI Advance 11中ClustalW progress;欧洲分子生物学开放软件系统EMBOSS 中的CHIPS和CUSP程序;系统进化树构建软件MEGA 5.0。
3.2.1密码子分析中涉及的指标
(1)有效密码子数(effective number of codons, ENC) (Wright, 1990);
(2) GC3s:基因中GC含量和密码子中不同位置的GC含量;
(3)密码子适应指数(codon adaptation index, CAI),密码子偏性达到最大值,即所有氨基酸均使用最优密码子,CAI值为1;而CAI值越小,密码子偏好程度就越低,相反偏好程度就越高;
(4) 同义密码子相对使用频率(relative synonymous codon usage, RSCU),指对于特定密码子在编码对应氨基酸相对同义密码子间的概率,密码子没有偏好时,该密码子的RSCU值等于1;RSCU值大于l,表明该密码子使用频率较高,反之使用频率就越低(Sharp and Li, 1986; 刘汉梅等, 2010)。
3.2.2基因密码子使用概率聚类分析
利用SPSS 13.0对9条LFY基因进行了基于密码子使用偏性的聚类。在对该基因密码子使用频率(RSCU)分析时,把每个基因作为一个对象,采用59个同义密码子(去除起始密码子和3个终止密码子)的RSCU值。基因间的距离分析利用密码子使用度的欧拉平方距离(Das et al., 2006),构建距离系数矩阵,并进行基于该系数的聚类分析。
作者贡献
张颖是本研究的实验设计和实验研究的执行人;樊秀彩,姜建福,孙海生完成数据分析,论文初稿的写作;李民,张永辉参与实验设计,试验结果分析;刘崇怀是项目的构思者及负责人,指导实验设计,数据分析,论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由现代农业产业技术体系建设专项资金(No. CARS-30-yz-1)资助。作者感谢两位匿名的同行评审人的评审建议和修改建议。
参考文献
Bradley D., Ratcliffe O., Vincent C., Carpenter R., and Coen E., 1997, Inflorescence commitment and architecture in Arabidopsis, Science, 275(5296): 80-83 http://dx.doi.org/10.1126/science.275.5296.80
Carbone A., Zinovyev A., and Képès F., 2003, Codon adaptation index as a measure of dominating codon bias, Bioinformatics, 19(16): 2005-2015 http://dx.doi.org/10.1093/bioinformatics/btg272
Carmona M.J., Cubas P., and Martinez-Zapater J.M., 2002, VFL, the grapevine FLORICAULA/LEAFY ortholog, is expressed in meristematic regions independently of their fate, Plant Physiology, 130: 68-77 http://dx.doi.org/10.1104/pp.002428
Das S., Paul S., and Dutta C., 2006, Synonymous codon usage in adenoviruses: influence of mutation, selection and protein hydropathy, Virus Res., 117(2): 227-236 http://dx.doi.org/10.1016/j.virusres.2005.10.007
Esumi T., Tao R., and Yonemori K., 2005, Isolation of LEAFY and TERMINAL FLOWER 1 homologues from six fruit tree species in the subfamily Maloideae of the Rosaceae, Sex. Plant. Reprod., 17: 277-287 http://dx.doi.org/10.1007/s00497-004-0239-3
Ikemura T., 1985, Codon usage and tRNA content in unicellular and multicellular organisms, Mol. Biol. Evol., 2(1): 13-34
Kanaya S.Y.Y., Kinouchi M., Kudo Y., and Ikemura T., 2001, Codon usage and tRNA genes in eukaryotes: correlation of codon usage diversity with translation efficiency and with CG-dinucleotide usage as assessed by multivariate analysis, J. Mol. Evol., 53(4-5): 290-298 http://dx.doi.org/10.1007/s002390010219
Liu H.M., Zhao Y., Gu Y., Zhang H.Y., and Huang Y.B., 2010, Characterization of codon usage of waxy genes in several plants, Journal of Nuclear Agricultural Sciences, 24(3): 476-481 (刘汉梅, 赵耀, 顾勇, 张怀渝, 黄玉碧, 2010, 几种植物waxy基因的密码子用法特性分析, 核农学报, 24(3): 476-481)
Murray E.E., Lotzer J., and Eberle M., 1989, Codon usage in plant genes, Nucleic Acids Res., 17(2): 477-498 http://dx.doi.org/10.1093/nar/17.2.477
Powell J.R., Sezzi E., Moriyama E.N., Gleason J.M., and Caccone A., 2003, Analysis of a shift in codon usage in Drosophila, J. Mol. Evol., 57(Suppl 1): S214-225 http://dx.doi.org/10.1007/s00239-003-0030-3
Ratcliffe O.J., Amaya I., Vincent C.A., Rothstein S., Carpenter R., Coen E.S., and Bradley D.J., 1998, A common mechanism controls the life cycle and architecture of plants, Development, 125(9): 1609-1615 http://dx.doi.org/10.1105/tpc.5.6.639
Shannon S., and Meeks-Wagner D.R., 1993, Genetic interactions that regulate inflorescence development in Arabidopsis, Plant Cell, 5: 639-655 http://dx.doi.org/10.1105/tpc.5.6.639 http://dx.doi.org/10.2307/3869807
Sharp P.M., and Li W.H., 1986, An evolutionary perspective on synonymous codon usage in unicellular organisms, J. Mol. Evol., 24(1-2): 28-38 http://dx.doi.org/10.1007/BF02099948
Wang H.C., and Hickey D.A., 2007., Rapid divergence of codon usage patterns within the rice genome, BMC Evol. Biol., 7(Suppl 1): S6 http://dx.doi.org/10.1186/1471-2148-7-14 http://dx.doi.org/10.1186/1471-2148-7-S1-S6 http://dx.doi.org/10.1186/1471-2148-7-6
Wong G.K.S., Wang J., Tao L., Tan J., Zhang J.G., Passey D.A., and Yu J., 2002, Compositional gradient s in gramineae genes, Genome Res., 12: 851-856 http://dx.doi.org/10.1101/gr.189102
Wright F., 1990, The 'effective number of codons' used in a gene, Gene, 87(1): 23-29 http://dx.doi.org/10.1016/0378-1119(90)90491-9
Wu X.M., Wu S.F., Ren D.M., Zhu Y.P., and He F.C., 2007, The analysis method and progress in the study of codon bias, Hereditas, 29(4): 7 (吴宪明, 吴松锋, 任大明, 朱云平, 贺福初, 2007, 密码子偏性的分析方法及相关研究进展, 遗传, 29(4): 7)
Zhang L., and Li W.H., 2004, Mammalian housekeeping genes evolve more slowly than tissue-specific genes, Mol. Biol. Evol., 21(2): 236-249 http://dx.doi.org/10.1093/molbev/msh010
Zhao W., Chen B.L., Wang M.Z., and Huang J.Z., 2010, Analysis on codon usage bias of fatty acid delta-6 desaturase genes from different algae, Journal of Tropical Oceanography, 29(3): 126-134 (赵薇, 陈必链, 王明兹, 黄建忠, 2010, 藻类△6脂肪酸脱饱和酶基因密码子偏好性分析, 热带海洋学报, 29(3): 126-134)