地方稻月亮谷花期控制基因Hd3a、Hd1、Edh1和Ghd7的多态性分析 
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2012 年, 第 10 卷, 第 23 篇 doi: 10.5376/mpb.cn.2012.10.0023
收稿日期: 2012年04月20日 接受日期: 2012年04月26日 发表日期: 2012年05月23日
引用格式(中文):
高东等, 2012, 地方稻月亮谷花期控制基因Hd3a、Hd1、Edh1和Ghd7的多态性分析, 分子植物育种(online) Vol.10 No.23 pp.1164-1170 (doi: 10.5376/mpb. cn.2012.10.0023)
引用格式(英文):
Gao et al., 2012, Analysis of Functional Nucleotide Polymorphisms in Hd3a, Hd1, Edh1 and Ghd7 Genes Controlling Heading Date in Yuelianggu Landrace, Fenzi Zhiwu Yuzhong (online) (Molecular Plant Breeding) Vol.10 No.23 pp.1164-1170 (doi: 10.5376/mpb.cn.2012.10.0023)
水稻抽穗期控制的关键基因已基本清楚,但是栽培稻花期多样性的分子机理仍然不清楚。本文利用重测序月亮谷的全基因组序列、Takahashi等(2009)在PNAS上发表的部分数据和Xue等(2008)在Nature genetics发表的部分数据,研究了Hd3a、Hd1、Edh1和Ghd7等4个基因的核苷酸多态性。结果显示,月亮谷Hd3a启动子区核苷酸多态性分布与前人报道的有所不同,发现5个特有的SNP位点,分别位于-1 851 bp、-1 720 bp、-700 bp、-681 bp和-1 607 bp处;1个特有的InDel,位于-523 bp处。月亮谷Hd1编码区其具有很高的多态性,发现6个特有的SNP/InDel位点,其中包括了所有4个移码位点,1个插入位点和1个缺失位点,月亮谷Hd1属无功能的等位基因类型。月亮谷Ehd1编码区SNP/InDel与前人研究结果相似。月亮谷Ghd7属于前人报道的有功能的等位基因Ghd7-1。月亮谷抽穗期相关基因复杂的多态性,一方面与其生长环境垂直海拔落差导致的“一山分四季,十里不同天”的复杂小生境有关,另一方面可能是其耐病、耐寒,适应性广的分子基础。
抽穗期是决定作物品种生育期及地区适应性的重要性状,受自身遗传因子和外界环境因素两方面决定,是开花基因时空顺序表达的结果(Yano et al., 2001)。水稻的全基因组序列测定已经完成(Goff et al., 2002; Yu et al., 2002),重测序项目发展迅速。序列数据库为侯选同源基因搜索提供了方便(Izawa et al., 2003)。SNP/InDel是一种在基因组中广泛存在的多态性(岳兵和邢永忠, 2005)。
在水稻上与农艺性状自然变异有关的功能核苷酸多态性(functional nucleotide polymorphisms, FNPs)已经得到了鉴定,其中包括花期相关基因Hd1、Edh1和Ghd7 (Yano et al., 2000; Doi et al., 2004; Xue et al., 2008; 薛为亚等, 2009)。根据GHD7蛋白序列可以把Ghd7分为5个等位基因,各等位基因间仅有几个氨基酸的不同,从而导致Ghd7的5个等位基因在地理分布上表现出明显的规律性。效应较大的Ghd7-1和Ghd7-3主要存在于栽种在亚热带、热带或夏季温度高且持续时间长的品种里;Ghd7-2分布于栽种在温带地区的粳稻品种里;完全缺失的Ghd7-0分布于华中和华南二季稻区的早稻品种里;蛋白质提前终止的Ghd7-0a则存在于东北黑龙江省的品种中(薛为亚等, 2009)。最近(Takahashi et al., 2009)等通过收集来自世界不同地区332份水稻品种(代表了水稻种质91%的多样性),来研究水稻花期的多样性,从开花时间上看所有收集的种质同样存在着极大的多样性,花期45~153 d不等(播种后到开花的时间)。同时在前人研究的基础上,选择涉及到影响水稻开花的相关基因来分析那些基因导致了这种开花时间的差异。研究发现Hd3a启动子序列的变异,Hd1核心编码序列的变异,Ehd1的表达量与对应的表型即开花时间的变异有着极显著的相关性。
本文以哀牢山哈尼梯田的传统代表品种(月亮谷)全基因组重测序为材料,借助目前在水稻花期基因中的研究结果,通过序列的搜索比对分析其SNP/InDel 的数量与分布;同时针对已报道的与开花适应性,花期多样性有关的基因的变异与类型与月亮谷的比较分析,为从分子水平上来解释当地传统品种能够持续种植上百年,从花期的角度提供一定的依据。
1结果分析
1.1 Hd3a启动子区的核酸多态性
Hd3a启动子区超过2 kb序列(包括5'末端非转录序列)的核苷酸多态性的分析结果显示(图1),月亮谷启动子区核苷酸多态性分布与前人报道的明显不同。核苷酸多态性变异介于两组(类型1, 类型2; 类型3, 类型4, 类型5, 类型6, 类型7)之间,与7种类型的任何一种都不相似,而且发现5个特有的SNP位点,分别位于-1 851 bp,-1 720 bp,-700 bp,-681 bp和-1 607 bp处;1个特有的InDel,位于-523 bp处。在启动子区的一些顺式作用元件(cis-elements)如CCAAT盒以及功能域如ARR没有核苷酸变异,这与文献报道一致。
.png) 图1 Hd3a启动子区核苷酸多态性 |
1.2 Hd1编码区的核酸多态性
Hd1编码区的核酸多态性 分析结果显示(图2),Hd1其具有很高的多态性。发现6个特有的SNP/InDel位点,其中包括了所有4个移码位点,1个插入位点和1个缺失位点。月亮谷Hd1基因包含有前人报道的871位基因位点上的移码突变,从而导致Hd1基因CCT结构域的部分失活,属无功能的等位基因类型。而对前人报道的非移码突变及提前终止位点,月亮谷Hd1基因类型介于上述类型之间;9311属于无功能的类型7,日本晴则属于有功能的类型5。
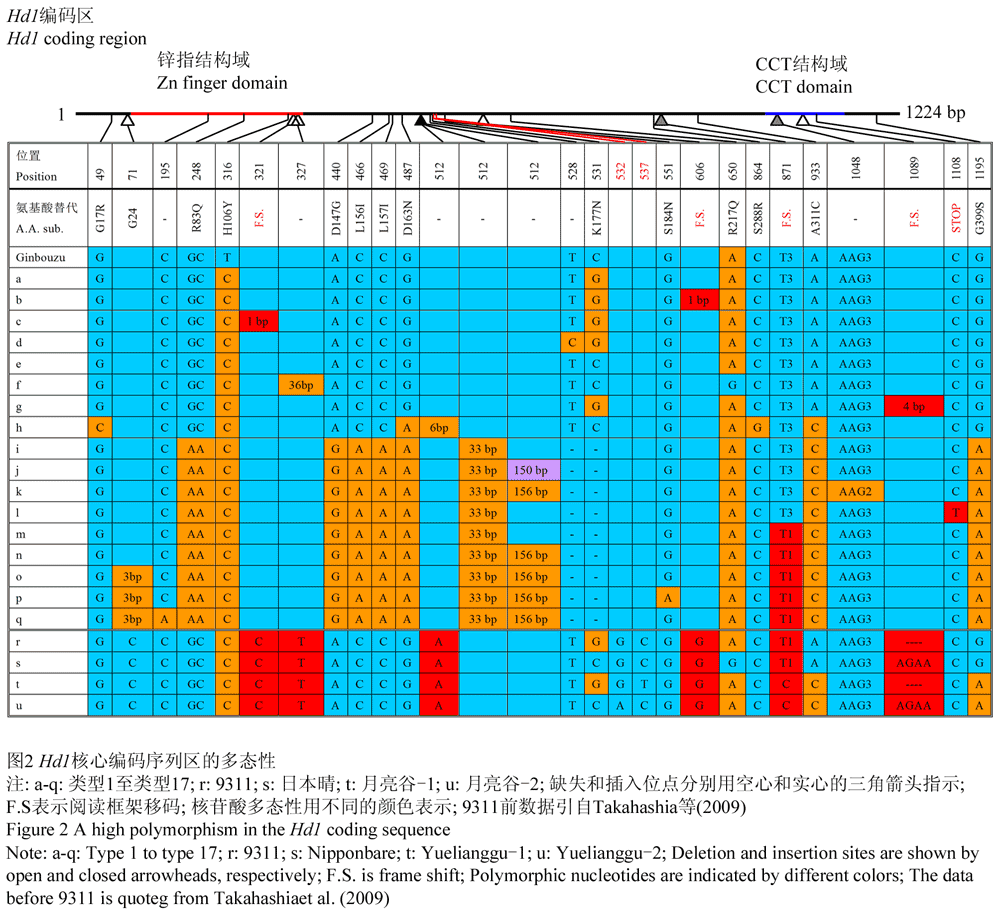 图2 Hd1核心编码序列区的多态性 |
1.3 Ehd1编码区的核酸多态性
Ehd1编码区的核酸多态性分析结果的显示(图3),月亮谷Ehd11编码区没有特有的SNP/InDel位点,与前人研究结果相似,可以归为类型3、类型4、类型5、类型6和类型7组成的组。
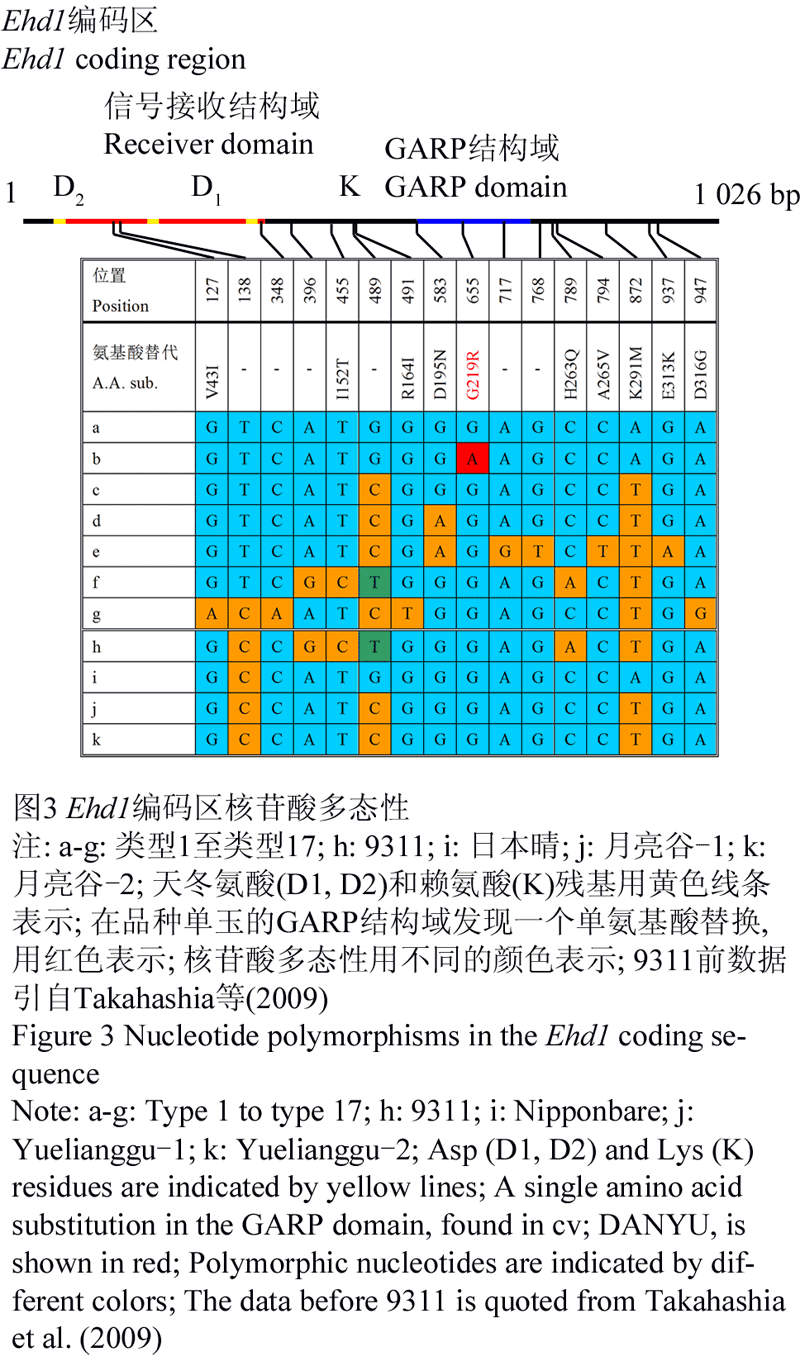 图3 Ehd1编码区核苷酸多态性 |
1.4 Ghd7等位基因的蛋白序列多态性
Ghd7是影响每穗粒数、抽穗期的一个数量性状位点(QTL),对水稻的一系列性状包括单穗粒数、株高和抽穗期都产生重要影响。通过对元阳传统水稻品种月亮谷Ghd7基因蛋白序列分析(图4),结果显示,月亮谷Ghd7属于前人报道的有功能的等位基因Ghd7-1。前人报道在野生稻种也发现具有功能的Ghd7-1,说明功能型的Ghd7-1可能是其他等位基因的祖先,随着水稻种植区域的扩大为了适应不同的地域环境,而衍生出其他等位基因,同时也导致开花时间对不同地区的适应性及多样性。
.png) 图4 Ghd7等位基因蛋白序列多态性 |
2讨论
植物对自然环境的适应一个很重要的机制就是通过对开花时间的调控进而在适当的季节开花,繁衍后代。水稻(Oryza sativa L.)从过去的8000至1万年前开始被驯化和繁育,有着很长的进化史(Doebley et al., 2006; Khush, 1997)。能够找到其适合的生态位和对自然环境的适应能力,是水稻生态繁荣的关键策略。水稻种植地理区域的扩展和广泛分布,产量的增加,一个很重要的因素就是水稻开花时间(花期)的多样性(Khush, 1997)。在特定环境下的开花时间调控,长期以来一直被理解为是一种很重要的适应性(Izawa, 2002)。
Takahashi等(2009)通过收集来自世界不同地区332份水稻品种(代表了水稻种质91%的多样性)来研究水稻花期的多样性,从开花时间上看所有收集的种质同样存在着极大的多样性,花期45 d~153 d不等(播种后到开花的时间)。同时在前人研究的基础上,选择涉及到影响水稻开花的相关基因来分析那些基因导致了这种开花时间的差异。研究发现Hd3a启动子序列的变异,Hd1核心编码序列的变异,Ehd1的表达量与对应的表型即开花时间的变异有着极显著的相关性。
Hd3a启动子2 kb的区域被认为是Hd3a表达所必须的,依据启动子以及5' UTR的核酸多态性分析,将收集的核心种可分为7种类型,对应于其开花时间多样性表型可将其分为两组;类型1类型2归为A组,类型3-7归为B组,表达分析表明B组Hd3a的表达显著高于A组,说明Hd3a启动子类型与开花时间有很显著的相关性。
月亮谷Hd3a启动子区的核酸多态性分布与前人报道的明显不同,介于A、B两组,与7种类型的任何一种都不相似,而且有5个特有的SNP位点。这些特征可能与哀牢山“一山分四季,十里不同天”的生态环境相适应,其相关性有待进一步研究。
Hd1编码区的核酸具有很高的多态性,可分为17个类型和15种独立的蛋白(Takahashi et al., 2009)。其中有9种无功能类型的核苷酸变异发生在CCT结构域中,CCT结构域编码一种核定位信号,在拟南芥中CO基因CCT的变异可导致CO蛋白的失活;相关性分析表明这种多态性与水稻花期有着显著的相关性;同时有功能的和无功能的Hd1基因类型具有明显的地区分布。月亮谷Hd1编码区的核酸较高的多态性,可能与哀牢山较高垂直海拔落差(144~2939.6 m)相关,其相关性有待进一步研究。
Xue等(2008)测序分析了东南亚的l9个品种,依据蛋白序列变异将Ghd7分为5个等位基因,蛋白质编码系列表明Ghd7-1与Ghd7-2有4个氨基酸的不同,导致Ghd7-2的功能较弱些;Ghd7-3与Ghd7-1和Ghd7-2有3个氨基酸的差别,同时有1个氨基酸变异是Ghd7-3特有的;Ghd7-0和Ghd7-0a是没功能的等位基因,Ghd7-0是功能域的删除而导致的,而Ghd7-0a是一个终止密码子的突变导致编码系列提前终止导致的。考察其地理分布,各等位基因具有明显的规律,效应较大的Ghd7-1和Ghd7-3主要分布在热带和亚热带以及夏季温度高且持续时间长的地区,Ghd7-2分布在温带粳稻品种里;Ghd7-0分布在华中和华南二季稻区的早稻里,Ghd7-0a则分布在夏季凉爽,水稻生育期短的东北黑龙江省的品种中(薛为亚等, 2009)。元阳传统水稻品种月亮谷持有功能的等位基因Ghd7-1,与其气候环境相符。但是,鉴于哀牢山的垂直海拔落差以及“一山分四季,十里不同天”的特征,有必要对哀牢山区不同小生境的月亮谷的Ghd7进行比较分析,极有可能发现其他类型的Ghd7。
3材料与方法
3.1哈尼族传统水稻品种月亮谷全基因组测序
哈尼族当地传统水稻品种月亮谷全基因组序列采取重测序的方法,我课题组出资,由华大基因(深圳)完成,并分别以9311和日本晴为模板拼接成2套月亮谷全基因组序列,分别记为月亮谷1 (9311)和月亮谷2 (日本晴)。
3.2基因序列的获取与比对
利用BIOEDIT (Kostka et al., 2008)将月亮谷的重测序序列建立本地BLAST数据库。然后用Hd3a、Hd1、Edh1和Ghd7的序列作查询序列搜索BLAST数据库发现基因片段,同时截取基因及上游2 kb的调控序列片段,序列比对用DNAMan软件进行,比对结果文件保存为.phy文件。依据Takahashi等(2009)发表的多态性位点进行汇总、做图。
作者贡献
高东是本研究的实验设计和实验研究的执行人;毛如志和魏富刚完成实验及数据分析;高东是论文的构思者及负责人,指导实验设计,数据分析,论文写作与修改。 全体作者都阅读并同意最终的文本。
致谢
本研究国家重点基础研究发展计划(973项目)课题(2011CB100406)资助。感谢唐有福在整个科研过程中提供的大量支持。
参考文献
Doebley J.F., Gaut B.S., Smith B.D., 2006, The molecular genetics of crop domestication, Cell, 127(7): 1309-1321
http://dx.doi.org/10.1016/j.cell.2006.12.006 PMid:17190597
Doi K., Izawa T., Fuse T., Yamanouchi U., Kubo T., Shimatani Z., Yano M., and Yoshimura1 A., 2004, Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1, Genes Dev., 18(8): 926-936
http://dx.doi.org/10.1101/gad.1189604
Goff S.A., Ricke D., Lan T.H., Presting G., Wang R., Dunn M., Glazebrook J., Sessions A., Oeller P., Varma H., Hadley D., Hutchison D., Martin C., Katagifi F., Lange B.M., Moughamer T., Xia Y., Budworth P., Zhong J., Miguel T., Paszkowski U., Zhang S., Colbert M., Sun W.L., Chen L., Cooper B., Park S., Wood T.C., Mao L., Quail P., Wing R., Dean R., Yu Y., Zharkikh A., Shen R., Sahasrabudhe S., Thomas A., Cannings R., Gutin A., Pruss D., Reid J., Tavtigaian S., Mitchell J., Eldredge G., Scholl T., Miller R.M., Bhatnagar S., Adey N., Rubano T., Tusneem N., Robinson R., Feldhaus J., Macalma T., Oliphant A., and Briggs S., 2002, A draft sequence of the rice genome (Oryza sativa L. ssp. japonica), Science, 296(5565): 92-100
http://dx.doi.org/10.1126/science.1068275 PMid:11935018
Izawa T., Oikawa T., Sugiyama N., Tanisaka T., Yano M., and Shimamoto K., 2002, Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice, Genes Dev., 16(15): 2006-2020
http://dx.doi.org/10.1101/gad.999202
Izawa T., Takahashi Y.J., and Yano M., 2003, Comparative biology comes into bloom: genomic and genetic comparison of flowering pathways in rice and Arabidopsis, Curr. Opin. Plant Biol., 6(2): 113-120
http://dx.doi.org/10.1016/S1369-5266(03)00014-1
Khush G.S., 1997, Origin, dispersal, cultivation and variation of rice, Plant Mol. Biol., 35(1-2): 25-34
http://dx.doi.org/10.1023/A:1005810616885 PMid:9291957
Kostka M., Uzlikova M., Cepicka I., and Flegr J., 2008, SlowFaster, a user-friendly program for slow-fast analysis and its application on phylogeny of Blastocystis, BMC Bioinformatics, 9: 341
http://dx.doi.org/10.1186/1471-2105-9-341 PMid:18702831 PMCid:2529323
Takahashi Y., Teshima K.M., Yokoi S., Innan H., and Shimamoto K., 2009, Variations in Hd1 proteins, Hd3a promoters, and Ehd1 expression levels contribute to diversity of flowering time in cultivated rice, Proc. Natl. Acad. Sci. USA., 106(11): 4555-4560
http://dx.doi.org/10.1073/pnas.0812092106 PMid:19246394 PMCid:2647979
Xue W., Xing Y., Weng X., Zhao Y., Tang W., Wang L., Zhou H., Yu S., Xu C., Li X., and Zhang Q., 2008, Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice, Nat. Genet., 40(6): 761-767
http://dx.doi.org/10.1038/ng.143 PMid:18454147
Xue W.Y., Xing Y.Z., Weng X.Y., Zhao Y., Tang W.J., Wang L., Zhou H.J., Yu S.B., Xu C.G., Li X.H., and Zhang Q.F., 2009, Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice, Zhongguo Jichu Kexue (China Basic Science), 2: 21-23 (薛为亚, 邢永忠, 翁小煜, 赵毓, 唐为江, 王磊, 周红菊, 余四斌, 徐才国, 李香花, 张启发, 2009, Ghd7自然变异是调控水稻抽穗期和产量潜 力的重要因素, 中国基础科学, 2: 21-23)
Yano M., Katayose Y., Ashikari M., Yamanouchi U., Monna L., Fuse T., Baba T., Yamamoto K., Umehara Y., Nagamura Y. and Sasaki T., 2000, Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS, The Plant Cell, 12(12):2473-2484
http://dx.doi.org/10.2307/3871242 PMid:11148291 PMCid:102231 http://dx.doi.org/10.1105/tpc.12.12.2473
Yano M., Kojima S., Takahashi Y., Lin H. and Sasaki T., 2001, Genetic control of flowering time in rice, a short-day plant, Plant Physiol., 127(4): 1425-1429
http://dx.doi.org/10.1104/pp.010710 PMid:11743085 PMCid:1540174
Yu J., Hu S., Wang J., Wong G.K., Li S., Liu B., Deng Y., Dai L., Zhou Y., Zhang X., Cao M., Liu J., Sun J., Tang J., Chen Y., Huang X., Lin W., Ye C., Tong W., Cong L., Geng J., Han Y., Li L., Li W., Hu G., Huang X., Li W., Li J., Liu Z., Li L., Liu J., Qi Q., Liu J., Li L., Li T., Wang X., Lu H., Wu T., Zhu M., Ni P., Han H., Dong W., Ren X., Feng X., Cui P., Li X., Wang H., Xu X., Zhai W., Xu Z., Zhang J., He S., Zhang J., Xu J., Zhang K., Zheng X., Dong J., Zeng W., Tao L., Ye J., Tan J., Ren X., Chen X., He J., Liu D., Tian W., Tian C., Xia H., Bao Q., Li G., Gao H., Cao T., Wang J., Zhao W., Li P., Chen W., Wang X., Zhang Y., Hu J., Wang J., Liu S., Yang J., Zhang G., Xiong Y., Li Z., Mao L., Zhou C., Zhu Z., Chen R., Hao B., Zheng W., Chen S., Guo W., Li G., Liu S., Tao M., Wang J., Zhu L., Yuan L., and Yang H., 2002, A draft sequence of the rice genome (Oryza sativa L. ssp. Indica), Science, 296(5565): 79-92
http://dx.doi.org/10.1126/science.1068037 PMid:11935017
Yue B., and Xing Y.Z., 2005, Progress on molecular and genetic studies of heading date in rice, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 3(2): 222-228 (岳兵, 邢永忠, 2005, 水稻抽穗期分子遗传研究进展, 分子植物育种, 3(2): 222-228)