植物落粒性状研究进展 
2. 海南省农作物分子育种重点实验室,三亚,572025
3. 东北林业大学盐碱地生物资源环境研究中心, 哈尔滨, 150040
 作者
作者  通讯作者
通讯作者
豆科基因组学与遗传学, 2011 年, 第 2 卷, 第 1 篇 doi: 10.5376/lgg.cn.2011.02.0001
收稿日期: 2010年12月05日 接受日期: 2010年12月24日 发表日期: 2010年12月27日
Luo et al., 2011, Research Progress of Seed-Shattering Habit in Plants, Douke Jiyinzuxue Yu Yichuanxue (online) (Legume Genomics and Genetics) Vol.2 No.1 pp.1-13 (doi: 10.5376/lgg.cn.2011.02.0001) (罗汝叶等, 2011, 植物落粒性状研究进展, 豆科基因组学与遗传学(online) Vol.2 No.1 pp.1-13 (doi: 10.5376/lgg.cn.2011.02.0001))
落粒性(shattering)是作物栽培和育种中的重要农艺性状,植物中落粒性状的驯化是人类文明史上重要成就之一。本文综述了双子叶及单子叶植物中的落粒性机制;以及影响落粒性的主要因素(形态学、解剖学特征,环境因素及遗传因素),并重点综述了这两类植物中落粒性的遗传机制,对今后的研究方向做了简要的展望。
植物的一个显著特征就是其器官(花器官、叶片、成熟或未成熟的果实、种子等)能够脱落。对于植物本身来说,器官的脱落是其生命过程中的重要生理现象,对物种的繁殖和传播有重要的生物学意义,但是很多植物(如小麦(Triticum aestivum L.)、水稻(Oryza sativa L.)、芝麻(Sesamum indicum L.)、大豆(Glycine max L.)、油菜(Brassica campestris L.)等)的果实脱落以及荚裂后种子的落粒在农业生产中会给农民造成严重的损失。
相对于野生品种来说,大多数作物的栽培品种对种子的脱落是高度抗性的,但是落粒(shattering)现象在一些植物中仍然存在着。如果延迟收割的话,会由于落粒而造成大量的损失。随着科学技术的快速发展,现在的农业生产中许多地方都是采用机械收割种子,但是机械收割会由于机械的物理影响造成严重的损失,所以通过增加种子的落粒抗性来减少损失是非常必要的。
单子叶植物与双子叶植物的果序以及每个果实的形状和结构都不相同。因此,许多人认为它们种子脱落的物理机制差异非常明显。然而,在单子叶植物及双子叶植物中落粒的发生都与离层的发育有关。因此,在落粒性机制多样化的不同品种中,可能存在解剖学、生理学以及遗传学方面存在着相似性(Kadkol et al., 1989)。近年来,越来越多的学者在不同的植物中对落粒性进行研究,如水稻(Oryza sativa L.) (Ji et al., 2010; Qin et al., 2010)、拟南芥(Arabidopsis thaliana L.) (Liljegren et al., 2004; Ogawa et al., 2009; Sorefan et al., 2009) 、油菜(Brassica campestris L.) (Kadkol, 2009)以及大豆(Glycine max L.) (Suzuki et al., 2010; Tsuchiya, 1987)等,并在解剖学特征及遗传学基础方面取得了一定的成果(Rajani and Sundaresan, 2001)。
1植物落粒抗性的进化
在许多植物中,落粒抗性已经成为驯化早期非常明显的结果。除人为的选择外,不落粒植物的种子被收获并重新播种,落粒抗性在驯化过程中能够自主进化。落粒抗性在野外环境中可能是一个适应性缺点,因此,物种进化后落粒性减少的植物被认为是依赖于人为的种植而生存。
在某些情况下,半落粒性在植物的生存中扮演着非常重要的角色。例如,在埃塞俄比亚草本植物燕麦和黑麦中,一些种子脱落并散播到地上,而留在植物上的种子被被收割并在下个季节中被作为栽培作物种植。这些植物在形态上模仿栽培作物,使得农民很难将它们从栽培作物中鉴定出来,因而适应了被收获和重新播种。这表明在一些物种中,落粒抗性是驯化的结果,而不是通过人类有目地选择的结果。对于一些具有很长栽培历史的牧草品种黑麦草属(Lolium0)和近属牛茅草(Festuca)和鸭茅(Dactylis glomerata)来说,尽管它们的落粒程度已经减少了并且种子保留水平也很高,但是其中只有少部分是通过人类有意识的育种和选择而来的。这可能为落粒抗性驯化过程中的自然选择提供了进一步的证据。目前水稻种植的目标不是种植不易落粒的或者容易落粒的品种,而最佳的落粒程度要借助于水稻育种中的标记辅助选择系统(MAS)来进行选择,现在普遍认为最佳的落粒程度是中度落粒。而大多数品种的落粒程度在一定程度上依赖于收获的形式(Qin et al., 2010)。
小麦祖先种子的脱落首先是通过穗轴的降解以及后来每个小穗分离而完成的。小麦野生品种和栽培品种间的一个重要的解剖学差异可能是由穗轴的脆性程度不同而产生的(Kadkol et al., 1989)。例如,在小麦二倍体栽培种(T. monococcum)中,穗轴是非常坚实的,只有在打谷的时候才会脱落。在它的野生品种(T. boeoticum)中,穗在成熟后降解并散播出种子。四倍体栽培种T. dicoccum和它的野生祖先T. dicoccoides之间也存在着相似的差异。小麦进化过程中的另一个重要的变化就是从具有顽强护颖的相对未开发的四倍体以及六倍体进化为自由脱粒的品种(Zohary, 1969)。已经有报道表明Q基因与六倍体以及某些四倍体小麦中的这一变化相关联(Mac Key, 1954; Muramatsu, 1986)。然而,Kerber和Rowland (1974)认为D基因组中出现了分离基因Tg控制植物自由脱粒的特性。
双子叶植物朝着不裂荚的方向进化和谷类作物朝着减少落粒性进化的趋势是相似的。例如,在一些豆科植物的进化中,豆荚果皮结构的改变、厚壁细胞层在数量上及方位上的大量变化以及中果皮木质化程度变低,这些转变已经导致了主动裂荚机制的丢失并且进化出不裂荚的品种。豆荚背部缝合线上维管束帽的厚度和长度以及豆荚壁的厚度与裂荚程度呈负相关,其在大豆进化过程中朝着裂荚程度减小的方向发展(Tiwari and Bhatia, 1995)。在十字花科的一些种(如油菜、拟南芥)中也具有相同的不裂荚的进化趋势,这些不裂荚植物果实的结构不形成离层组织。
2落粒性机制
植物的落粒性状导致收获过程中大量的损失,基本上在所有植物中都普遍存在着落粒性状。然而,其在不同的植物(如双子叶植物及单子叶植物中)中发生机理可能不同,在植物中所表现的相关特征也各不相同。最初的植物进化过程中,落粒性状对种子的散播起了非常重要的作用,其对物种的生存至关重要。因此,自然选择使果实的解剖学及形态学朝落粒性及荚裂性方向发展。
2.1脱落及开裂
落粒性的一个普遍特征就是植物的个别种子或者单位种子(小麦及大麦的小穗)能够从植株上脱落,或长角果(油菜的长荚果)以及大豆的豆荚裂开散播出种子。这种果实脱落和开裂是借助于离层细胞发育的机械性减弱进行的。就像在谷类及草本植物中一样,离层在果实及穗轴的附着区发生;在豆荚及长荚果植物中,离层是在果实活瓣的连接区产生的。
Burson等(1983)对草本本植物Paspalum notatum、P. dilatatum、Panicum colpratum和P. maximum进行研究,表明了落粒过程中离层的个体发生和作用。离层在孕穗期早期的小穗与穗轴的附着区域开始发育,并且在孕穗期的最后阶段经过开花期而发育完全。开花期13 d后,离层的中间薄层裂解,细胞分散以及维管束断裂引起了脱落组织中空隙的形成,导致穗的脱落。然而,在一些禾本科植物中,落粒性状似乎不依赖于脱落组织的发育。例如,在虉水草(Phalaris aquatica)中,穗轴镶嵌在两个护颖压缩后的基底间,护颖发育并且干燥变硬,导致小穗轴的断裂以致小穗的脱落,而在小穗轴中没有脱落组织(McWilliam, 1980)。
由于油菜角果的中央出现了隔膜(replum),隔膜将果皮包裹着两个小室分开,果皮边缘与隔膜连接的地方发育成裂荚区(Dehiscence zone DZ)。裂荚区的细胞最后开始退化并且弱化了果皮间、瓣间以及隔膜间的结合。细胞的凝聚力的损失仅在裂荚区发生并且是由于中间薄层的降解引起的(Spence et al., 1996)。
2.2主动裂荚机制
双子叶植物的角果失水后通过主动或者被动的裂荚机制而开裂。主动裂荚是角果失水后由一个内置机制产生的压力最终导致裂荚的过程,这个过程的发生不需要借助于外界的力量。而在被动裂荚中,离层组织的发育增加了果实脱落性状的发生,这完全是由外部力量的影响而不是内置机制引起的裂荚(如果皮上不产生拉力)。
在拟南芥中,裂荚机制主要分为四个主要的步骤(Rajani and Sundaresan, 2001)。(1)由SHP1/SHP2基因控制的长角果瓣边缘(valve margin)的裂荚区在长角果受精之后开始分化。(2)果实脱水后瓣膜层内部与裂荚区发生木质化、细胞层木质化并且瓣的外层细胞收缩使得果实产生拉力。(3) ALC基因在瓣膜边缘与假隔膜之间启动裂荚区细胞的一个子集(如非木质化细胞(NLC)层)分化,随后形成一个非木质化的边界。因为在alc突变体中裂荚区细胞的木质化程度没有发生改变,所以木质化与非木质化细胞层的分化可能是两个独立发生的事件。在alc突变体的相同位置细胞发生生态型木质化,如果要阻止非木质化细胞层细胞的木质化,研究细胞的特征要比研究细胞的位置更加重要。(4)非木质化细胞层最后经过自溶和细胞壁的降解(这些降解可能是经过细胞壁降解酶而引起的)。当这些细胞降解之后,果实中存在的拉力辅助瓣膜从假隔膜上分离开来使得种子散播。
在主动裂荚中,果皮拉力的产生是由于果实瓣的厚壁细胞在特定的方位上分化的结果。在蚕豆及羽扇豆中,外果皮由厚壁细胞组成,在方向上与内果皮的厚壁细胞层相反。这两个果皮在方向上都与荚的众轴线成45°角。然而,与细胞轴相关的微纤维的方位在内果皮及外果皮中都是相同的,因为在这两个果皮层中,细胞轴的方位不一样。所以,干燥后这两个细胞层在方位上的最大收缩程度不一样。不同的收缩使豆荚产生拉力,这个拉力防止瓣附着于脱落层。瓣弯曲并且爆炸性地裂开(Fahn and Zohary, 1955)。在大豆(Monsi, 1942)、亚麻籽(Holden, 1956)及芝麻(Ashri et al., 1964)中可能存在着相同的机制。Christiansen等(2002)对大豆豆荚的在成熟过程中的不同阶段的背部和腹部的缝合线横切面进行显微镜观察,发现大豆的裂荚区(dehiscence zone, DZ)和十字花科裂荚区功能相当。这表明豆科作物的裂荚机制可能与十字花科作物的裂荚机制相似。
3影响落粒性的相关因素
落粒性是很多作物都具有的一个重要特征,在最初的生长过程中,落粒性使种子可以分散到环境中,保证种子物种的繁殖,但是在生产中如果不能及时收获种子,会导致大量的种子丢失,给种子生产带来一定的困难。然而,导致落粒的因素很多,目前普遍认为主要有下列三个因素:(1)植物果序及荚果的解剖结构;(2)环境因素;(3)遗传因素。
3.1与落粒抗性相关的解剖学及形态学特征
许多作物中落粒抗性的变化被认为是与果实中果序落粒区的解剖学特征及植物中的某些形态学特征的变化相联系。这些解剖学特征通常有助于落粒区变坚实,促使落粒减少。双子叶植物和单子叶植物由于果实的形态结构不相同,所以它们与落粒性相关的解剖学特征可能也不相同。
3.1.1单子叶植物落粒性相关的的解剖学特征
大多数现代的栽培作物(除水稻外)的品种的穗柄处没有产生离层,因此,大都对落粒具有高度的抗性(McWilliam, 1980)。而在水稻中,离层仍然存在,甚至在抗落粒的水稻栽培品种中仍然具有一个不完全发育的离层。而在不同品种中离层的形态各不相同。也有一些不落粒的品种不含有离层,在易落粒的品种中,离层在果皮与维管束之间发育而成(Ji et al., 2006) (图1)。落粒的抗性程度可能与维管束及离层间厚壁组织的厚度呈正相关。在Zizania aquatica野生品种中,穗轴中的离层促使其脱落,但是落粒抗性的发生是由于离层细胞不能够溶解与分裂而产生的(Hanten, 1975)。
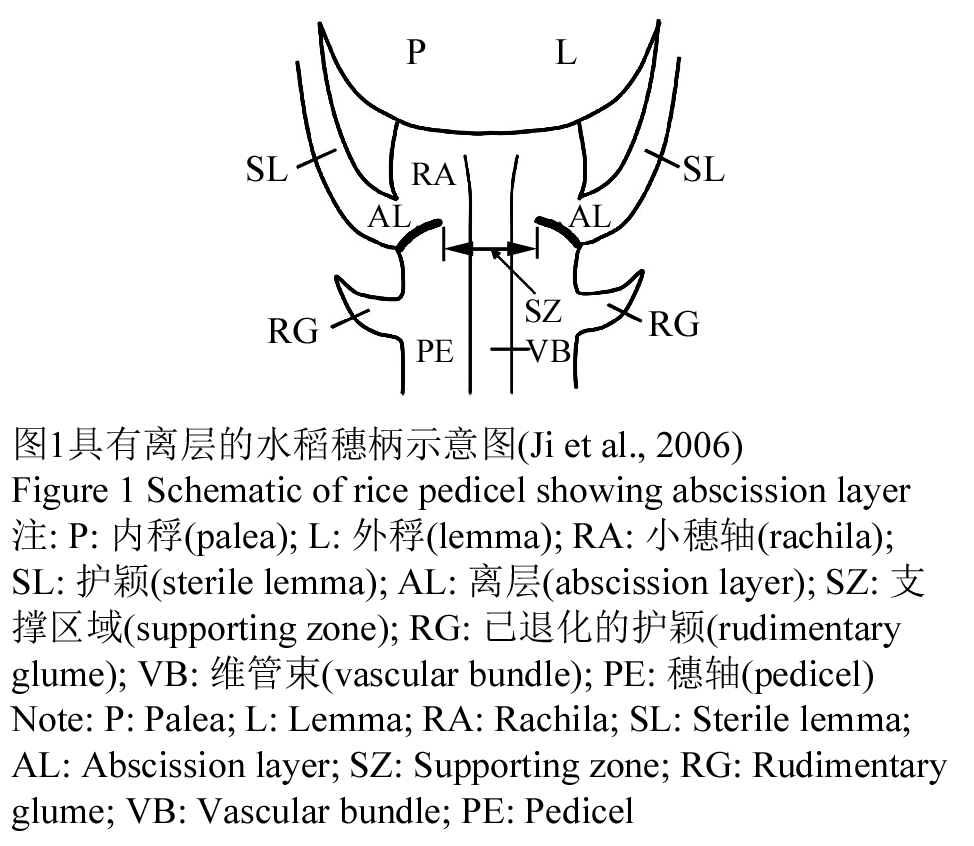 图1 具有离层的水稻穗柄示意图(Ji et al., 2006) 注: P: 内稃(palea); L: 外稃(lemma); RA: 小穗轴(rachila); SL: 护颖(sterile lemma); AL: 离层(abscission layer); SZ: 支撑区域(supporting zone); RG: 已退化的护颖(rudimentary glume); VB: 维管束(vascular bundle); PE: 穗轴(pedicel) Figure 1 Schematic of rice pedicel showing abscission layer Note: P: Palea; L: Lemma; RA: Rachila; SL: Sterile lemma; AL: Abscission layer; SZ: Supporting zone; RG: Rudimentary glume; VB: Vascular bundle; PE: Pedicel |
水稻落粒性被认为是由位于护颖和花梗间的接合处离层(abscission layer, AL)的形成以及后来的中间薄层和细胞壁的降解而引起的(Patterson, 2001; Roberts et al., 2002)。离层大约是在抽穗前16~20 d幼穗为5~30 mm时形成的(Jin, 1986),当种子成熟后,离层的细胞降解,增加了谷粒从母体中脱落的机会。和落粒程度呈反比的穗轴抗张强度(BTS)在抽穗后10~20 d后减弱(Ji et al., 2010)。Qin等(2010)等用一个来自于容易落粒的品种‘Samgang’和中度落粒的品种‘Nagdong’杂交后的双单倍体系(doubled ha- ploid line, DHL)种群来观察了落粒及不落粒品种的果实中的离层的发育(图2)。结果在‘Samgang’中出现了离层,在‘Nagdong’的穗柄组织中观察到部分离层。他们认为落粒性不仅仅是由离层控制的,离层在穗柄组织中所处的区域和位置只是影响落粒程度的因素之一。
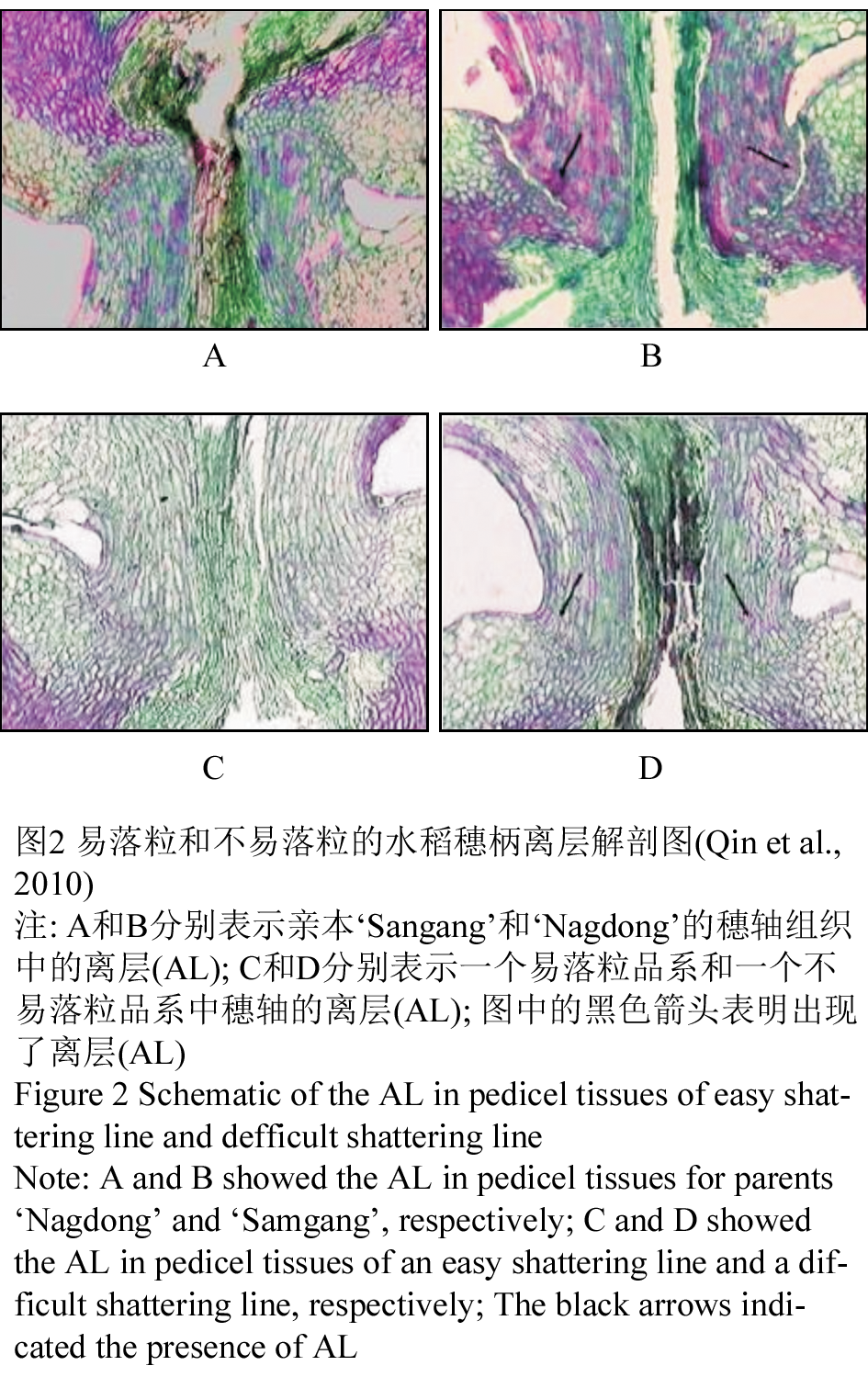 图2 易落粒和不易落粒的水稻穗柄离层解剖图(Qin et al., 2010) 注: A和B分别表示亲本‘Sangang’和‘Nagdong’的穗轴组织中的离层(AL); C和D分别表示一个易落粒品系和一个不易落粒品系中穗轴的离层(AL); 图中的黑色箭头表明出现了离层(AL) Figure 2 Schematic of the AL in pedicel tissues of easy shattering line and defficult shattering line Note: A and B showed the AL in pedicel tissues for parents ‘Nagdong’ and ‘Samgang’, respectively; C and D showed the AL in pedicel tissues of an easy shattering line and a difficult shattering line, respectively; The black arrows indicated the presence of AL |
3.1.2双子叶植物落粒性相关的解剖学特征
在一些豆科植物及其它双子叶植物中,同时存在着离层和主动裂荚机制,落粒抗性是与位于缝合线处离层的损失或主动裂荚机制相联系的,或者与两者都有关系。
Tsuchiya等(1987)在大豆(Glycine max L.)中对落粒的以及抗落粒性的品种的豆荚形状和大小与落粒性状之间的关系进行研究。发现除了抗性品种中豆荚厚度/豆荚宽度比易裂荚的品种大之外,其他因素(如豆荚长度、宽度、厚度、弯曲度、豆荚壁的厚度等)在这两个品种中没有多大的区别。Calson (1973)认为直接引起裂荚的原因是由于豆荚内部厚壁层的细胞水分丢失后产生的拉力。在百脉根(Lotus corni- culatus)中,豆荚的解剖学特征对裂荚起了非常重要的作用。其裂荚被认为主要是由果皮中细胞方位的改变(如豆荚的不等位膨胀和收缩),以及中果皮的少量木质化而引起的(Repkova and Hofbauer, 2009)。
油菜果实和拟南芥具有相似的结构,都是由两个融合的雌蕊叶组成的雌蕊受精后发育成为长角果(Bowman et al., 1999)。B. napus的荚果由两个瓣组成,在其发育末期,荚果的两个辦由一个条带连接,长角果完全成熟后,两个瓣向两个方向伸展(Child et al., 2003)。甘蓝型油菜(Brassica napus)荚果的开裂与裂荚区中充满胶质的中间薄片的退化有关。裂荚是因为薄的荚皮存在一个裂荚区域,此区域的部分细胞分开,最后由于拉力的作用使荚果开裂(Child et al., 2003)。Summer等认为抗裂荚能力很可能与荚皮结构或裂荚区域相关而不是与荚的大小相关(Summers et al., 2003)。Kadkol等(1986)报道了油菜中离层是由荚果薄壁和非木质化的细胞构成,存在于易裂荚品种的长角果缝合线处,而在抗裂荚的品种中没有离层的存在(图3)。
图3 B. napus荚果的结构特征(Agius et al., unpublished) Figure 3 Structural features of a B. napus pod |
Wagstaff等将拟南芥的长角果壁的发育分为4个阶段:第一:绿色角果的成熟阶段(开花期后10 d);第二:50%的角果变黄阶段(开花期后20 d);第三:100%的角果变黄阶段(开花期后22 d);第四:角果完全成熟并在裂荚点脱水阶段(开花期后24 d)。通过显微镜观察发现,长角果壁或瓣包含有三个由外部和内部表皮联系的叶肉层,但是与内表皮相邻的是一层特殊的细胞,他们具有非常厚的壁,然后分化成为内果皮。当角果完全伸长后,内表皮细胞开始降解,并且内果皮的细胞壁已经开始变厚。内表皮细胞的分解和死亡持续到开花期后22天(第三阶段),内表皮层只出现在两层紧贴的细胞壁上,不具有可见的细胞质内容物。内果皮持续加厚到第四阶段,也就是在裂荚之前,然后细胞壁层出现裂开的现象(Wagstaff et al., 2009)。拟南芥的两个瓣(valve)包裹着种子并在内部沿着角果与假隔膜(replum)相连,当果实发育后假隔膜向外发展,果实成熟后瓣从假隔膜上分开散播出种子(Dinneny et al., 2005)。长角果裂荚是在一系列与瓣边缘形成的裂荚区相关的协调事件下发生的(Rajani and Sundaresan, 2001)。裂荚区(Dehiscence zone, DZ)是瓣与假隔膜分离的边缘区域,由瓣边缘的离层与木质化层组成(图4)。离层(sepa- ration layer)是夹在假隔膜侧边的相对较大的薄层细胞层与一组在瓣边缘的木质化细胞间的一条不稳定的非木质化细胞条带(Ferrandiz, 2002)。瓣边缘的木质化层是与瓣木质化的内部亚表皮层相连接,在成熟后长角果中产生拉力使瓣从假隔膜上和离层裂开,离层的中间薄层通过水解酶使细胞壁的支持力减弱而被分开(Spence et al., 1996)。目前的研究表明拟南芥中的裂荚区(DZ)在长角果裂开的过程中起着非常重要的作用,同在离层的发育以及内部亚表皮层的转录网络中也起着非常重要的作用(Tan et al., 2009)。
在芝麻中,落粒抗性可能与缝合线处的中果皮的发育相关。
图4 假隔膜(replum)与瓣(valve)区域的横切面示意图(Rajani and Sundaresan, 2001) 注: Ex: 外果皮; Ms: 中果皮; Ena, Enb:内果皮层;V: 假隔膜维管系统; DZ: 在假隔膜和瓣的边界线处形成的裂荚区 Figure 4 Schematic representation of transverse section at the replum valve region Note: The exocarp (Ex), mesocarp (Ms), the two endocarp layers Ena and Enb, and the replum vasculature (V) are shown. The mardehiscence zone (DZ) forms at the valve replum boundary |
3.1.3影响落粒性的形态学特征
人们在一些植物中已经发现落粒抗性与植物的形态特征相关(如果序的密度以及主干上的主要枝条的角度) ((Kadkol et al., 1989))。在小麦中,落粒抗性与外颖的木质化以及果序的紧凑性相关联(Chang, 1943)。
油菜中的落粒抗性被认为与长角果中瓣的厚度相关(Rai et al., 1959)。而Jonsson等(1968)在油菜中观察到落粒抗性与植物的某种形态特征间只有很低的相关性。Kadkol等(1984)报道了大量的田间落粒与主干的枝条角度间的正相关性,但是与另一些植物特征(如高度、长角果附着于果序的轴线间的角度、主要枝条的数目、枝条上每个长度单位上长角果的数目、长角果的长度以及长角果梗的长度)并没有表现出明显的相关性。
综上所述,在单子叶植物中,落粒可能与离层的发育相关;而在双子叶植物中,植物的裂荚与荚果的瓣边缘的裂荚区相关。而植物的很多形态学特征与植物的落粒性状只表现出很低的相关性,甚至有的特征对落粒性状并没有严重的影响。
3.2影响落粒性的环境因素
在豆科作物以及一些其它的双子叶植物中,有报道表明落粒性状与一些环境因素相关;而在单子叶植物及少数双子叶植物中,有关这方面的报道非常少,落粒性更多地是由遗传因素决定的。
豆科植物的裂荚不是在荚果一开始形成就出现的,而是发生在种子的收获期,荚果发育到一定的程度后,在一定的环境条件下发生的。百脉根(Birdsfoot trefoil)的克隆B74的荚果在相对湿度为29.5%时开始出现裂荚,裂荚与相对湿度、水分平衡是相互依赖的;在相对较低的湿度条件下,果荚皮含水量会迅速下降到裂荚临界点(Metcalfe te al., 1957)。Tsuchiya (1987)认为尽管还没有鉴定出影响裂荚的任何特定环境因子,但是低湿度,高温,温度、湿度以及干燥的快速改变等都可能减少裂荚的发生。他还认为裂荚是由环境因素及遗传因素共同决定的,并且环境因素中豆荚中的水分含量是最主要的因素。Metcalfe等(1957)也认为在豆科作物中相对湿度是影响裂荚的主要环境因素。
3.3遗传因素
落粒性状在单子叶植物及双子叶植物中都是主要由遗传因素决定的,其遗传性是物种在长期进化过程中形成的稳定的遗传特性,是对自然环境的一种适应机制。植物将种子散播于环境中,保证了起物种的迅速繁殖。Elmoneim (1993)对野豌豆(Vida sativa L.)的遗传性研究发现,野生品种中的普通野豌豆的不落粒性状是由一个简单的隐性基因控制的,而在栽培品种中的落粒性状是由一对显性的等位基因控制。Grant (1996)认为在莲花中(Lotus)落粒抗性是一个高遗传性特征并且是由一个以上的基因控制的。
尽管没有明确的比例,但是Woods和Clark (1976)在野生水稻(Z. aquatica)中观察到易落粒性相对于抗落粒性状是显性的并且F2代的数据表明其是由简单的遗传控制的,可能是由一个或者两个基因控制。Elliot和Perlinger (1977)在抗落粒品种和野生的易落粒落粒品种杂交的F2代中观察到3:1与9:7的比例,并且易落粒性相对于抗落粒性是显性的。他们认为有两个互补基因影响着水稻的落粒性状。
在大豆中,有报道表明裂荚性状相对于不裂荚性状是显性的,可能是由四个基因控制的。抗裂荚品种与农艺学特征(如开花期和种子蛋白含量)之间有很低的负相关性,并且抗裂荚的广义遗传值为89%~98% (Caviness, 1969),Tsuchiya等(1987)认为裂荚是由环境因素及遗传因素决定的,并且在环境因素中,豆荚的水分含量最为重要。同时估计出裂荚性状的遗传性平均为93%,用最大可能性的方法对9个抗裂荚与敏感的杂交品系中的8个进行评估,并用Castle-Wright的方法对所有杂交品系评估,发现裂荚性状是由1~2个基因决定的。Tukamuhabwa等(2000)认为大豆中的裂荚是由两对基因控制的并且裂荚性状相对抗裂性状荚表现出部分显性。
4落粒性的遗传机制
早期的植物落粒性的研究都侧重于生理生化、环境影响等方面,对落粒性的遗传性方面的研究比较少。近年来,随着分子遗传学及细胞生物学的发展,很多科学家开始研究脱落层的发育及裂荚的机制方面的问题,并侧重于研究控制脱落层以及开裂遗传基础以及这控制裂荚的基因的调控网络。
4.1单子叶植物落粒抗性的遗传机制
4.1.1水稻
从野生水稻品种驯化出不易落粒品种的水稻栽培过程中,人们选择出了控制理想谷粒脱落程度的等位基因,但是直到最近几年,随着分子生物学及细胞遗传学的飞速发展,关于落粒性状的遗传机制才有了突破性的进展。
2006年,Konishi等(2006)对易落粒籼稻品种Kasalath和不易落粒的粳稻品种Nipponbare杂交的F2代群体的落粒性状的QTL进行分析,在第一染色体长臂上发现了一个贡献率为68.6%的主要位点qSH1 (seed shattering in chromosome 1)。qSH1的5’端调控区域中的一个SNP (single nucleotide polymer- phism)导致离层不能够形成从而使作物不发生落粒现象。同年,Li等(2006)用籼稻品种和一年生野生稻作为亲本得到F2群体,对水稻中的落粒性状的QTL进行遗传分析,发现了一个贡献率为69%的主效QTL位点(SH4),并且为显性遗传,并最终定位于水稻第4染色体的1.7 kb范围内。这个基因编码一个未知功能的转录因子,控制水稻的离区发育。随后的亚细胞定位结果(定位在细胞核中)也认为SH4是一个转录因子。转基因实验表明栽培种与野生型的水稻落粒性只因第1个外显子的单核苷酸改变而使栽培种中的赖氨酸代替了野生种中的天冬氨酸。另一个能够影响水稻落粒性状的单一基因SHA1 (Shattering 1),编码植物中thihelix家族的特异性转录因子。被定位到第4染色体上的包含有一个开放阅读框的5.5 kb的基因片段上。其编码的氨基酸与SH4基因编码的氨基酸有98%的序列是一致的,而且是等位基因。后来在YJCWR品种的突变体中发现,其cDNA的+237的位置上的G转变为了T,导致氨基酸的第79个赖氨酸变成了天门冬氨酸,这使得落粒性状的丢失。SHA1基因与SH4基因不同的是,它并不能影响水稻种离层的形成,但是可能与离层的细胞壁的降解有关(Lin et al., 2007)。
另一个与水稻落粒性相关的基因SH-H,位于第7染色体上的34 kb的区域上。此基因被称为Oryza sativa CTD phosphatase like 1 (Ji et al., 2010),编码一个含有一个保守的羧基端结构域(CTD)的磷酸酶区域,亚细胞定位及生物化学分析表明OsCPL1蛋白是一个核磷酸,后生动物中的CTD磷酸酶的一个普遍特征就是与细胞的分化有关。结果表明OsCPL1基因在穗的发育期间抑制离层的分化,从而使水稻不能够脱落。但是水稻中的离层不是由单一的基因控制的,并且落粒性不仅仅由离层决定的;从物理力的角度来说,离层在穗组织中的区域和位置只是影响谷类作物落粒程度的因素之一(Qin et al., 2010)。谷粒的脱落是可能是由一个复杂的遗传调控网络控制,并受很多因素的影响。所以还需要进一步的研究来更清楚地阐明这个调控网络。
4.1.2其它作物
Srinivas对不同倍性以及不同叶耳形态的小麦穗轴的脱落进行研究发现穗轴的脆性是由位于A、B和D基因组上的许多基因组控制的。对穗轴的脱落起主要作用的Q位点控制穗轴中离层的发育,离层的厚度依赖于这个位点上隐性的等位基因q的量。与普通小麦的落粒抗性相关的颖壳韧度是由位于染色体2D的长臂上一个Tg基因控制的,具有顽强颖的品种相对于自由脱粒的来说是显性的。Porter (1959)认为Wichita×Cimarron杂交的小麦的落粒抗性是由多基因中的隐性基因控制的。在Wichita×Blackhull杂交中,落粒抗性是由两对主要基因以及修饰器控制的。
Chapman和Hockett (1976)认为在大麦中落粒性是显性基因控制的。并且有证据表明是由至少两对基因控制的。
4.2双子叶植物落粒抗性的遗传机制
4.2.1大豆
大豆是世界上重要的粮食、油料作物,目前中国大豆生产总量不能满足国内对大豆的需求,其主要原因是大豆产量低(唐晓飞等, 2010)。大豆的裂荚性状是影响其产量的因素之一。而目前国内外对大豆裂荚性状的研究大多是对其QTL的分析。
Suzuki等(2009)检查了近等基因系(near-isogenic lines NILs)中控制裂荚的主要质量性状位点(QTL),揭示出这个抗裂荚的QTL的潜在影响机制。他们还在这些NILs中发现这些品系的豆荚长度、宽度以及长度方面都没有表现出不同。进行豆荚的解剖学分析发现大多数裂荚在背部的缝合线处开始,这在NILs中也没有变现出明显的不同。因此,他们认为qPDH1控制裂荚的过程中并没有明显地改变豆荚的形态学结构。
2002年,Christiansen等(2002)对大豆豆荚的成熟过程中不同阶段的背部和腹部的缝合线横切面进行显微镜观察,发现大豆的裂荚区(dehiscence zone, DZ)和十字花科裂荚区在功能上相似。酶分析发现豆荚中出现了内切-1,4-β-葡聚糖(endo-1,4-β-glu- canases)和内切聚半乳糖醛酸酶(endopolygalacturo- nases),这两个酶在豆荚裂荚区积累并在成熟时达到最大值。通过PCR克隆出一个编码内切聚半乳糖醛酸酶的单一的不完整的cDNA,并将这个克隆用来进行提取编码内切聚半乳糖醛酸酶的完整基因的探索。在植物转录载体pCAMBIA1301中的uidA (GUS)基因的上游克隆出大约有1.2 kb的5’端上游序列,并转录进拟南芥中。表达分析发现内切聚半乳糖醛酸酶主要在大豆中裂荚相关组织中出现并且与裂荚前中间薄层细胞的断裂有关。在十字花科植物中,长角果的开裂依赖于长角果壁的降解及特殊离层(dehiscence zone)的断开,离层的降解是两个瓣边缘的分开。而离层的分离依赖于细胞壁修饰酶(大多数都是内切聚半乳糖醛酸酶, endo-PG, 被称为RDPG1)来促使中间薄层的断裂(Petersen et al., 1996)。虽然在大豆中的SDPG与拟南芥中的ADPG以及油菜中的RDPG的相似性只有43%和57%,并且表达模式也不相同,但是其在大豆中和果实及种子的离层细胞壁的降解相关。
2005年,Funatsuki等(2005)用简单序列重复(SSR)标记对一个重组自交体系(RIL)种群的裂荚性状进行QTL分析,复合区间作图发现一个主要QTL (qPDH1)位于连锁群(LG)J的SSR标记的Sat-093与Sat-366之间。并在裂荚品种与抗裂荚的RIL (重组自交品系)的杂交F2代种群中确定此QTL的位置和影响。估计这些标记间的距离为2.9 cm。这可以用于大豆育种中的分子辅助选择(MAS)。2008年,他们在四个不同的遗传背景下,再次证明了qPDH1是位于Sat-093与Sat-366之间的,并且证明这个主要的QTL在大豆中控制着裂荚性状(Funatsuki et al., 2008)。
2009年,Yamada等(2009)对几个种群的DNA标记进行遗传分析,在来自于易裂荚品系Toyomu- sume以及抗裂荚品系Harosoy的F2代种群中,一个与裂荚相关的主要QTL被定位于qPDH1附近,这与F4:5代群体中所定位的一样;Wasekogane和Kariyutaka作为抗裂荚的亲本的杂交F2代种群的QTL也位于qPDH1附近。所以他们认为尽管抗裂荚基因来自于不同的遗传背景,但是这个主要的QTL仍然被定位于qPDH1。在QTL处,杂合基因型表现出很高的裂荚性,说明裂荚抗性性状表现为近隐性性状,并且很可能位于qPDH1处。
2010年