研究报告
蒺藜苜蓿PEPC蛋白生物信息学分析及在碱胁迫中功能的预测 
 作者
作者  通讯作者
通讯作者
豆科基因组学与遗传学, 2016 年, 第 7 卷, 第 6 篇
收稿日期: 2016年08月22日 接受日期: 2016年08月22日 发表日期: 2016年08月22日
引用格式(中文):
任永晶, 宋婷婷, 姜柳, 许慧慧, 才华, 2016, 蒺藜苜蓿PEPC蛋白生物信息学分析及在碱胁迫中功能的预测, 分子植物育种, 14(2): 344-351
引用格式(英文):
Ren Y.J., Song T.T., Jiang L., Xu H.H., and Cai H., Bioinformatics analysis of PEPC proteins from Medicago truncatula and functional predictions under alkaline stress, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 14(2): 344-351
磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是一种广泛存在于自然界中的代谢酶,在高等植物中参与光合作用的碳固定、生物合成前体的供应、pH的调控等多种生物学过程。本研究通过生物信息学方法,识别、筛选、获得蒺藜苜蓿(Medicago truncatula)PEPC(MtPEPC)的氨基酸序列Medtr2g076670.2,并对与Medtr2g076670.2相似的25个豆科PEPC基因进行系统进化树分析。着重对4个MtPEPCs和6个GmPEPCs基因进行功能域分析、蛋白二级结构域预测及组织表达特异性分析,以推测蒺藜苜蓿PEPC基因的功能。结果表明,该10个基因分为两个Group分支,两个分支中存在不同的亚细胞定位信号,并且两个分支中大豆基因在地上和地下组织之间存在差异。通过蛋白相互作用的预测分析,PEPC蛋白可与苹果酸脱氢酶(Malate dehydrogenase, MDH)、丙酮酸激酶(Pyruvate kinase, PK)及2个未知蛋白存在相互作用;在碱胁迫下,GsPEPCs基因与苹果酸脱氢酶基因是“共表达”基因。可以推测,PEPC对碱胁迫的反应,可能是通过调节有机酸含量实现的。通过对MtPEPC蛋白和GmPEPC蛋白的生物信息学分析,获得了其相应的分子生物学特征,为PEPC蛋白在碱胁迫反应中的功能提供理论依据。
蒺藜苜蓿(Medicago truncatula)是一种一年生的二倍体苜蓿,作为一种优良的牧草在全世界范围内被广泛种植。蒺藜苜蓿具有基因组小、生长期短、自花授粉、根瘤固氮、遗传转化效率高等特点,已成为豆科生物学以及逆境响应研究的模式植物(李紫薇等, 2014; 陈爱民等, 2006; 宋辉等, 2014; 姜格格等, 2013)。大豆与蒺藜苜蓿同属于豆科植物,在遗传上有较高的相似性,大豆基因组及转录组方面的研究可为蒺藜苜蓿、紫花苜蓿、豌豆、三叶草等豆科植物的研究提供更多的参考(刘志敏等, 2015)。
磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)是一种广泛存在于自然界中的代谢酶,分布在古菌,细菌,藻类和维管植物中。在高等植物中参与光合作用的碳固定、生物合成前体的供应、对pH的调控及豆科作物根瘤固氮过程信号级联反应(Xu et al., 2007)。有研究报道,在植物面对干旱(周宝元等, 2011; 方立锋, 2008)、强光(李霞等, 2005)、盐(Echevarría et al., 2001)、低磷胁迫(Wu et al., 2003)和冷害(Jane, 2011)等逆境情况下,PEPC蛋白的活性有所提高。由此表明,PEPC在植物非生物胁迫反应中也发挥一定的作用。然而,PEPC在碱胁迫反应中的作用尚无明确的报道。本研究以MtPEPC蛋白序列Medtr2g076670.2为目的基因,借助蒺藜苜蓿基因组数据库、大豆基因组数据库及公共数据库(NCBI, GEO, Phytozome),对蒺藜苜蓿PEPC蛋白及与其同源的大豆PEPC蛋白Glyma.12G229400.1进行生物信息学分析,根据大豆PEPC蛋白的结构域、组织表达差异、碱胁迫表达谱及蛋白质相互作用预测蒺藜苜蓿PEPC蛋白在碱胁迫反应中的生物学功能,为蒺藜苜蓿该类基因的功能研究提供参考。
1结果与分析
1.1序列获取与功能域分析
以蒺藜苜蓿基因Medtr2g076670.2为种子序列,共获得具有相似保守结构域的苜蓿PEPC蛋白4个,大豆PEPC蛋白6个(表1)。苜蓿PEPC基因分别定位于第2、8、4号染色体上,大豆PEPC基因分别定位于第6、12、13号染色体上。
.png) 表1 蒺藜苜蓿和大豆PEPC蛋白的获取与定位 |
1.2结构功能域分析与同源序列比对
蒺藜苜蓿Medtr2g076670.2在161~966处为由806个氨基酸组成的PEPcase结构域(图1);与之最相近的大豆蛋白为Glyma.12G229400.1 (GmPEPC1),相似性为94.30%,也具有完整的PEPcase结构域,由此推测这两个基因可能具有相似的生物学功能。
.png) 图1 蒺藜苜蓿和大豆PEPC结构域 |
1.3蛋白质序列系统进化树分析
根据Medtr2g076670.2的氨基酸序列,在Phytozome数据库中检索6个豆科物种中其他的同源蛋白,经过筛选得到含有完整PEPC的结构域的氨基酸序列25个,利用MEGA5.0软件并采用NJ法构建系统进化树(图2)。25个PEPC蛋白一共被分成了3个分支,Group I分支中,蒺藜苜蓿蛋白Medtr2g076670.2、Medtr8g463920.2 (“■”所示)与大豆Glyma.12G229400.1 (“●”所示)相似性最高,Glyma.12G161300.1、Glyma.06G229900.1也属于此分支;在Group II中,Medtr2g092930.1与大豆中的Glyma.12G210600.1、Glyma.13G290700.1两个蛋白相似性最高,而Medtr4g079860.1则与大豆的Glyma.06G277500.1具有更高的相似性,并与四季豆的PEPC蛋白聚为一组;在GroupIII中,黄瓜、草莓和苹果中的几个蛋白聚为一组。此6个大豆PEPC基因在非生物胁迫反应中的作用均未见报道,并且均不是根瘤增强型的基因(Hata et al., 1998)。但值得注意的是,已经报道盐胁迫响应的野生大豆GsPEPC和GsPEPC1 (Qi et al, 2014)与大豆Glyma.12G161300.1、Glyma.06G229900.1具有98%~99%的相似性。由此暗示,PEPC蛋白在参与基本的生物代谢外,还通过其它的生物学过程响应非生物胁迫。
.png) 图2 Medtr2g076670.2基因系统进化树 |
1.4 PEPC蛋白二级结构分析
信号肽预测显示,蒺藜苜蓿和大豆PEPC蛋白都没有信号肽序列和跨膜区;均含有Ser/Thr和Tyr蛋白激酶磷酸化活性位点;并且蛋白主要由α–螺旋、不规则卷曲和延伸链组成。然而亚细胞定位预测发现,GroupI中与GroupII中的蛋白亚细胞定位信号存在差异,蒺藜苜蓿和大豆表现同样的特性。GroupI中蒺藜苜蓿和大豆PEPC蛋白主要定位于核、微体、叶绿体类囊体膜、叶绿体基质、线粒体(仅Medtr2g076670.2无线粒体定位信号)等几个组织,而GroupII中的5个蛋白却没有叶绿体类囊体基质的定位信号。亚细胞定位信号的差异暗示,同是PEPC类蛋白,在不同的组织部位和亚细胞结构中,参与不同的功能;并且,二级结构的差异决定了PEPC蛋白具有多种生物学功能(表2)。
.png) 表2 蒺藜苜蓿和大豆同源蛋白一、二级结构预测信息 |
1.5 PEPC蛋白组织特异性分析
由于蒺藜苜蓿数据库中没有基因表达谱数据,4个MtPEPCs基因序列及功能域与6个GmPEPCs基因具有较高的相似性,借助GmPEPCs基因组织特异性分析数据推测MtPEPCs基因的功能。根据进化树的分支,分别获得GroupI分支的3个PEPC蛋白(Glyma.12G229400.1, Glyma.12G161300.1, Glyma.06G229900.1)和GroupII分支的3个PEPC蛋白(Glyma.12G210600.1, Glyma.13G290700.1, Glyma.06G277500.1)根、根毛、叶片、根瘤、花等9个组织表达特性图谱(图3)。由此可见,GroupI分支的3个GmPEPCs基因在地下组织(根毛、根中及根瘤)中表达量高于叶片,茎等地上组织;相反GroupII分支的GmPEPCs基因主要在地上组织中表达。基因组织表达的特性与定位信号的分析、系统进化树分析具有类似的结果。PEPCs基因的生物学功能在地上部位和地下组织中存在不同的功能分工。
|
注: a: Group I分支中3个GmPEPCs基因组织表达分析 (Glyma.12G229400.1, Glyma.12G161300.1, Glyma.06G229900.1); b: Group I分支中3个GmPEPCs基因组织表达分析(Glyma.12G210600.1, Glyma.13G290700.1, Glyma.06G277500.1) |
.png)
1.6蛋白质相互作用分析
同样借助大豆PEPCs蛋白的相互作用,辅助了解MtPEPCs蛋白的功能和调控机制。利用STRING交互式数据库搜索与Glyma.12G229400.1蛋白相互作用的蛋白,构建蛋白相互作网络(图4)。与GmPEPC1(Glyma.12G229400.1, Glyma.12G35840.1)互作的10个蛋白中(表3),有5个是苹果酸脱氢酶(Malate dehydrogenase, MDH)。MDH可以催化苹果酸与草酰乙酸间的可逆转换,主要参与TCA循环、光合作用、C4循环等代谢途径(汪新颖等, 2009)。其中Glyma.11G04720.1具有乙醛酸循环体活性,说明GmPEPC蛋白与乙醛酸循环代谢也存在关系。Glyma.20G02980.1、Glyma.07G35110.3这两个蛋白都有丙酮酸激酶活性,丙酮酸激酶参与糖酵解途径,是糖无氧酵解途径的三个关键调节点之一。丙酮酸激酶在第三阶段的最后一步中催化磷酸烯醇式丙酮酸(PEP)转变为丙酮酸,产生能量ATP。在机体缺氧时糖酵解途径能迅速提供能量,为细胞的生命活动提供部分能量,其生成的一些中间产物是脂类和氨基酸等合成的前体(欧艳, 2010)。根据与GmPEPC1蛋白互作的蛋白类型可以看出PEPC参与TCA循环、光合作用、C4循环、乙醛酸循环、糖酵解等多种代谢途径。
.png) |
|
表3 互做蛋白的注释名称和参与的生物学过程 |

1.7碱胁迫基因表达分析
从NCBI的GEO中下载了NaHCO3胁迫下大豆根(GSE17883)和叶(GSE20323)中的基因表达数据,分析了4个大豆PEPC基因及与之相互作用的9个蛋白在碱胁迫中的表达情况(图5),推测MtPEPCs基因在碱胁迫下可能的生物学功能。在NaHCO3胁迫下同一个基因在地上和地下组织中表达量及表达模式并不相同。Glyma.12G229400.1在根中的表达量明显高于在叶中的表达量,与之互作的蛋白Glyma17G10880.1在根中的表达量与在叶中的表达量也明显不同。除Glyma.12G229400.1基因外,Glyma06G277500.1在根和叶中的表达量都比较低,并且在碱胁迫下差异变化较少;Glyma12G161300.1在根和叶中的表达量均较低,为下调表达;Glyma12G210600.1在根和叶中的表达量均较高,但是在根中,该基因为上调表达,而在叶中则为下调表达。另外,根据基因表达聚类发现,在根和叶中,Glyma.10G34150.1和Glyma.08G06820.1基因表达模式与Glyma.12G229400.1相似,可确定该3个基因为“共表达”基因。该结果与相互作用网络构建的结果一致,说明GmPEPC1蛋白与该2个蛋白存在相互关系,Glyma.10G34150.1为未知蛋白,而Glyma.08G06820.1注释为苹果酸脱氢酶。由此推测,PEPC蛋白参与苹果酸脱氢酶的代谢途径,并通过该途径代谢的变化响应碱胁迫。
分别对大豆PEPC蛋白根和叶中的基因表达模式进行聚类分析(图6),可以发现在叶中,3 h时基因的表达量达到了最大值,而在根中基因表达变化差异最大的时间点为6 h。但是也不难看出,根是最先响应碱胁迫的,胁迫1 h后,基因上调表达到最高值。
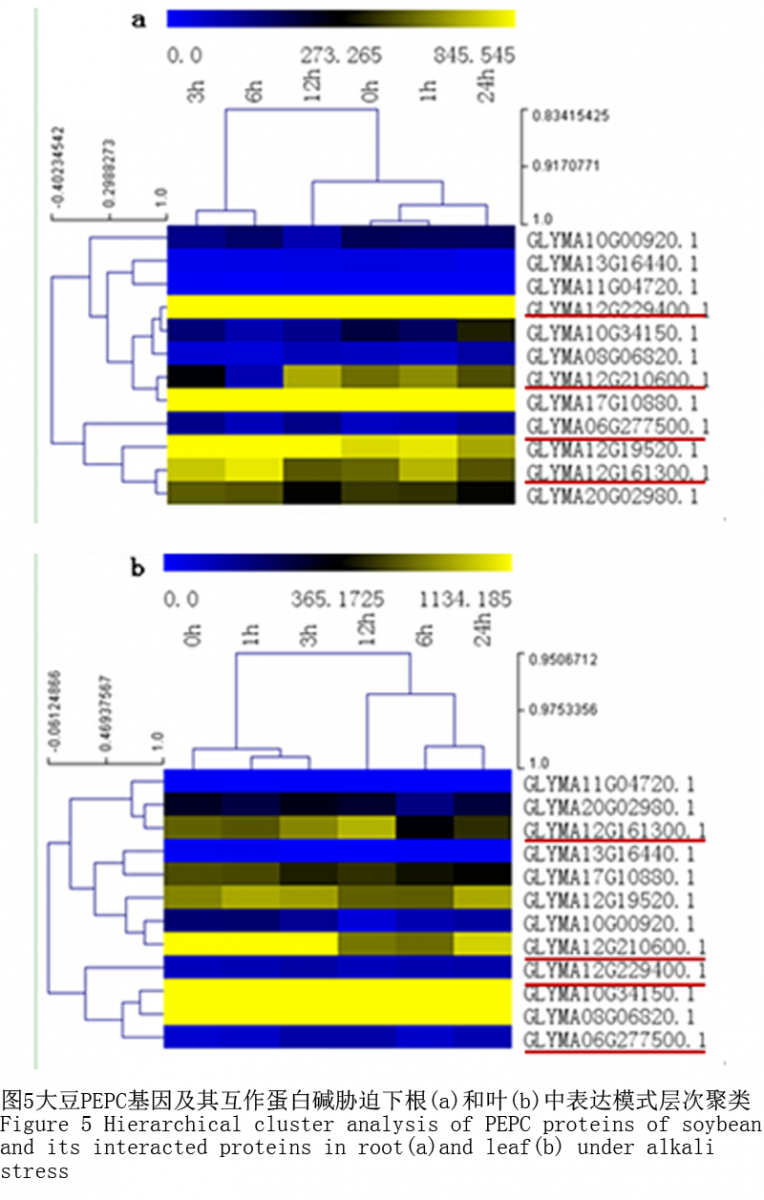 |
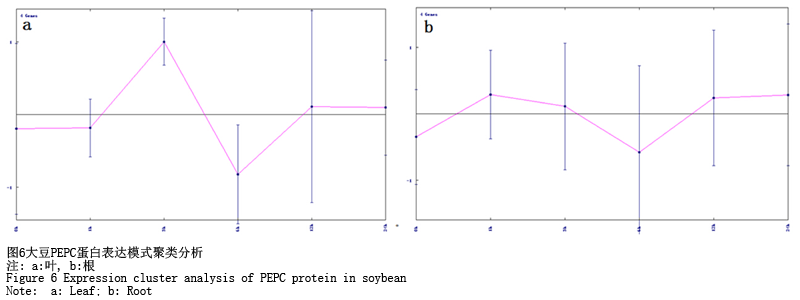 图6 大豆PEPC蛋白表达模式聚类分析 注: a:叶, b:根 |
2讨论
据报道,PEPC基因有不同的类型。在高等植物中PEPC有2个已知的功能形式,光合型的PEPC在C4光合作用和景天酸代谢(CAM)途径中催化大气CO2初始固定形成C4二羧酸(Deng, et al., 2015);非光型的PEPC已在C3植物的叶子和各种器官中发现,如:根发育中的种子和豆科植物根瘤中等。在根瘤中,PEPC发挥共生固氮的作用,在碳氮代谢中起到重要的作用(Naima et al., 1997; Nomura et al., 2006)。本研究发现与Glyma.12G229400.1相似的25个PEPC蛋白一共被分成了3个分支,其中Group I和Group II 2个分支中的PEPC蛋白在亚细胞定位信号存在着差异。除此之外,Group I的3个GmPEPCs基因在地下组织(根毛,根中及根瘤)中表达量高于叶片,茎等地上组织;而Group II分支的GmPEPCs基因主要在地上组织中表达。根据碱胁迫表达谱数据分析的结果,根最先响应碱胁迫,胁迫1 h后,基因上调表达达到最高值。综上所述,PEPCs基因对非生物胁迫的反应除了要考虑其不同类型外,还应考虑地上和地下2个不同的组织区域,地下部分首先感受胁迫信号,其势必与地上组织存在不同的功能分工。
Hata (1998)等发现,GmPEPC7基因与GmPEPC1/15及GmPEPC4基因在编码区具有98%~99%的相似性,但是3'-UTR的相似性仅有37%。而正是由于其非编码区的差异,导致GmPEPC7基因是根瘤增强型的PEPC,另两个基因则为持家基因。本文中所述的6个大豆PEPC基因均不是根瘤增强型的基因,但是不可忽视其非编码区及其启动子的差异。蒺藜苜蓿中4个PEPC基因在氨基酸序列上与大豆6个PEPC基因具有91%~98%的相似性,蒺藜苜蓿中PEPC基因是否具有与其相似的大豆PEPC基因的功能,还不能单纯从其序列的相似性推测,还需要考虑其非编码区和启动子顺式作用元件的差异。
根据pH稳态调节假说,高浓度HCO3–能激活PEPC,具有最佳活性,其结果是,产生苹果酸防止大量胞浆碱化。如果苹果酸的产生使胞浆过度酸化那么PEP的活性被抑制,而NADP-苹果酸酶(NADP-ME)被激活,增加苹果酸脱羧酶酶活以防止细胞质酸化(Sakano et al., 1998)。蛋白互作分析显示,与PEPC蛋白相互作用的9个蛋白中,有5个蛋白为苹果酸脱氢酶;另外,根据基因表达聚类发现,该2个基因为“共表达”基因,由此可以推定,PEPC可能是通过调控苹果酸脱氢酶的活性响应植物耐碱。另外,在根中,根瘤的发育也伴随着苹果酸的产生和分泌,这个过程的调控也与PEPC蛋白相关。
3材料与方法
3.1序列获取与定位
利用NCBI网站(http://www.ncbi.nlm.nih.gov/genbank/),搜索根据序列的相似性搜索蒺藜苜蓿MtPEPC (Medtr2g076670.2)的同源蛋白。在Phytozome(http://www.phytozome.net/)中搜索与Medtr2g076670.2蛋白具有相同结构域的大豆同源蛋白。
3.2结构功能域分析及同源基因系统进化树构建
运用在线分析工具SMART(http://smart.embl-heidelberg.de/)进行结构功能域分析,删除不含有完整PEPC保守结构域的蛋白序列,只保留存在完整PEPC保守结构域的并与Medtr2g076670.2具有80%以上相似性的蛋白。采用邻近相连算法(Neighbor-Joining),利用MEGA5.0软件构建系统进化树。
3.4蒺藜苜蓿及大豆PEPC蛋白的二级结构预测
蛋白质序列信号肽及跨膜结构域预测分别采用CBS (http://www.cbs.dtu.dk/services/SignalP/)服务器中蛋白预测工具signal peptide和TMHMM Serverv.2.0。亲疏水性分析(http://www.cbs.dtu.dk/services/NetPhos/)、磷酸化位点分析(http://www.expasy.eh/egi-bin/protscal.pl)及二级结构的预测(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat. pl?page=/NPSA/npsa_hnn.html)均利用在线工具进行分析。
3.6蛋白质相互作用分析
应用STRING交互式数据库(http://string-db.org/)搜索与Glyma.12G229400.1(Glyma.12G35840.1,PPC1)蛋白相互作用的其他蛋白质信息,搜索条件限制在10个蛋白内,构建的蛋白相互作网络。
3.7 PEPC基因及与其相互作用蛋白基因碱胁迫表达差异分析
在NCBI的GEO(http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE9730)数据库中下载大豆的根和叶在碱(NaHCO3)胁迫下的表达谱数据,利用MeV4.9软件进行表达模式和层次聚类分析。
作者贡献
才华指导实验设计,进行论文写作与修改。任永晶是本研究实验工作的具体执行人,完成数据分析和论文初稿的写作。宋婷婷、姜柳和和许慧慧完成基因表达谱数据、进化树分析及数据整理、文章校稿工作。全体作者都阅读并同意最终的文本。
致谢
本研究由国家自然科学青年基金(31302022)和黑龙江省自然科学基金(C2015018)共同资助。感谢哈尔滨工程大学王洋和鲍佩华博士对本文序列分析方法的指导。
Chen A.M., Lian R.L., Sun J., and Wang Y.Z., 2006, Leguminous model plant-Medicago truncatula, Zhiwu Shenglixue Tongxun (Plant Physiology Communications), 42(5): 997-1003 (陈爱民, 连瑞丽, 孙杰, 王彦章, 2006, 豆科模式植物—蒺藜苜蓿, 植物生理学通讯, 42(5): 997-1003)
Echevarría C., Garcia-Mauriño S., and Alvarez R., 2001, Salt stress increases the Ca2+-independent phosphoenolpyruvate carboxylase kinase activity in Sorghum leaves, Planta, 214(2): 283-287
Fang L.F., 2008, Physiological impact of PEPC gene overexpressed rice plants under drought stress, Thesis for M.S., Chinese Academy of Agricultural Sciences, Supervisor: Zhao M., pp.06-01 (方立锋, 2008, 转PEPC基因水稻干旱逆境下的生理反应, 硕士学位论文, 中国农业科学院, 导师: 赵明, pp.06-01)
Jane A.L., 2011, C4 Cycles: Past, Present, and Future Research on C4 Photosynthesis, Plant Cell, 23(11): 3879–3892 Hata S., Izui K., and Kouchi H., 1998, Expression of a soybean nodule-enhanced phosphoenolpyruvate carboxylase gene that shows striking similarity to another gene for a house-keeping isoform, The Plant Journal, 13(2): 267-273
Deng H., Zhang L.S., Zhang G.Q., Zheng B.Q., Liu Z.J., and Wang Y., 2015, Evolutionary history of PEPC genes in green plants: Implications for the evolution of CAM in orchids, Molecular Phylogenetics and Evolution, available online, doi:10.1016/j.ympev.2015.10.007
Jiang G.G., Song L.L., Guo D.L., Cai H.S., Guo C.H., and Shu Y.J., 2013, Genome-wide association mapping of aluminum tolerance in Medicago truncatula,, Caoye Xuebao (Acta Prataculturae Sinica),22(4):170-178 (姜格格, 宋丽莉, 郭东林, 蔡洪生, 郭长虹, 束永俊, 2013, 蒺藜苜蓿耐酸铝性状的全基因组关联分析, 草业学报, 22(4): 170-178)
Li X, Jiao D.M., and Dai C.C., 2005, The Response to Photooxidation in Leaves of PEPC Transgenic Rice Plant(Oryza sativa L.), Zuowu Xuebao (Acta Agronomica Sinica), 31(4): 408-413 (李霞, 焦德茂, 戴传超, 2005, 转PEPC基因水稻对光氧化逆境的响应, 作物学报, 31(4): 408-413)
Li Z.W., Ma T., Liang G.T., and Guo J., 2014, Photosynthesis in Leaves of Medicago truncatula under Salt Stress, Xibei Zhiwu Xuebao (Acta Botanica Boreali-occidentalia Sinica), 34(10): 2070-2077 (李紫薇, 马天, 梁国婷, 国静, 2014, 蒺藜苜蓿叶片光合作用对盐胁迫的响应, 西北植物学报, 34(10): 2070-2077)
Liu Z.M., Liu W.X., Jia X.T., Zhang Z.S., and Wang Y.R., 2015, Genome-wide analysis of LEA gene family in Medicago truncatula, Caoye Kexue (Pratacultural Science), 32(3): 382-391 (刘志敏, 刘文献, 贾喜涛, 张正社, 王彦荣, 2015, 蒺藜苜蓿LEA基因家族全基因组分析, 草业科学, 32(3): 382-391)
Nomura M., Mai H.T., Fujii M., Hata S., Izui K., and Tajima S., 2006, Phosphoenolpyruvate carboxylase plays a crucial role in limiting nitrogen fixation in Lotus japonicus nodules, Plant and Cell Physiol., 47(5): 613-621
Ou Y., 2010, Study on the mechanism of the change of the conformation of pyruvate kinase in the activation of ion and substrate, Thesis for M.S., FuDan University, Supervisor: Yu S.N., pp.05-10 (欧艳, 2010, 丙酮酸激酶在活化离子和底物诱导下的构象变化机理研究, 硕士学位论文, 复旦大学, 导师: 余绍宁, pp.05-10)
Sakano K., 1998, Revision of biochemical pH-stat: involvement of alternative pathway metabolisms, Plant Cell Physiol., 39(5):467-473
Song H, and Nan Z.B., 2014, Genome-wide identification and analysis of WRKY transcription factors in Medicago truncatula, Yichuan (Hereditas (Beijing)), 36(2): 152-168 (宋辉, 南志标, 2014, 蒺藜苜蓿全基因组中WRKY转录因子的鉴定与分析, 遗传, 36(2): 152-168)
Naima B., Jean L. P., Eliane D., Jean P. R., Martine A. D., Jean V., Pierre G. and Raymond C.,1993, 1997,Regulatory phosphorylation of C4 Phosphenolpyruvate carboxylase.Plant Physiol.101:891-897
Wang X.Y., Wang B.,Hou S.T., and Zhu G.P., 2009, Structure and function of malic dehydrogenase, Shengwuxue Zazhi (Jounal of Biology), 26(4): 69-72 (汪新颖, 王波, 侯松涛, 朱国萍, 2009, 苹果酸脱氢酶的结构及功能, 生物学杂志, 26(4): 69-72)
Wu P, Ma L. G., Hou X. L., Wang M. Y., Wu Y.R., Liu F.Y., and Deng X.W., 2003, Phosphate starvation triggers distinct alterations of genome expression in Arabidopsis roots and leaves, Plant Physiology, 132(3):1260-1271
Qi X., Li M., Xie M., Liu X., Ni M., Shao G., Song C., Yim A. K., Tao Y., Wong F., Isobe S., Wong C., Wong K., Xu C., Li C., Wang Y., Guan R., Sun F., Fan G., Xiao Z., Zhou F., Phang T., Liu X., Tong S., Chan T. Yiu S., Tabata S., Wang J., Xu X., and Lam H., 2014, Identification of a novel salt tolerance gene in wild soybean by whole-genome sequencing, Nature Communications, 5
Xu W., Sato S.J., Clemente T.E., and Chollet R., 2007, The PEP-carboxylase kinase gene family in Glycine max (GmPpcK1–4): an indepth molecular analysis with nodulated, non-transgenic and transgenic plants, The Plant Journal, 49(5): 910-923
Zhou B.Y., Ding Z.S. and Zhao M., 2011, Alleviation of drought stress inhibition on photosynthesis by overexpression of PEPC gene in rice, Zuowu Xuebao (Acta Agronomica Sinica), 37(1): 112-118 (周宝元, 丁在松, 赵明, 2011, PEPC过表达可以减轻干旱胁迫对水稻光合的抑制作用, 作物学报, 37(1): 112-118)